

Cold-Dependent Expression and Alternative Splicing of Arabidopsis Long Non-coding RNAs
- PMID: 30891054
- PMCID: PMC6413719
- DOI: 10.3389/fpls.2019.00235
Cold-Dependent Expression and Alternative Splicing of Arabidopsis Long Non-coding RNAs
Abstract
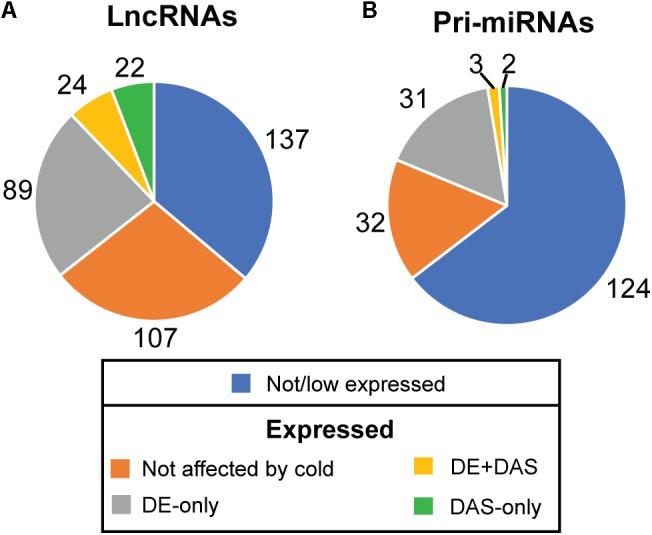
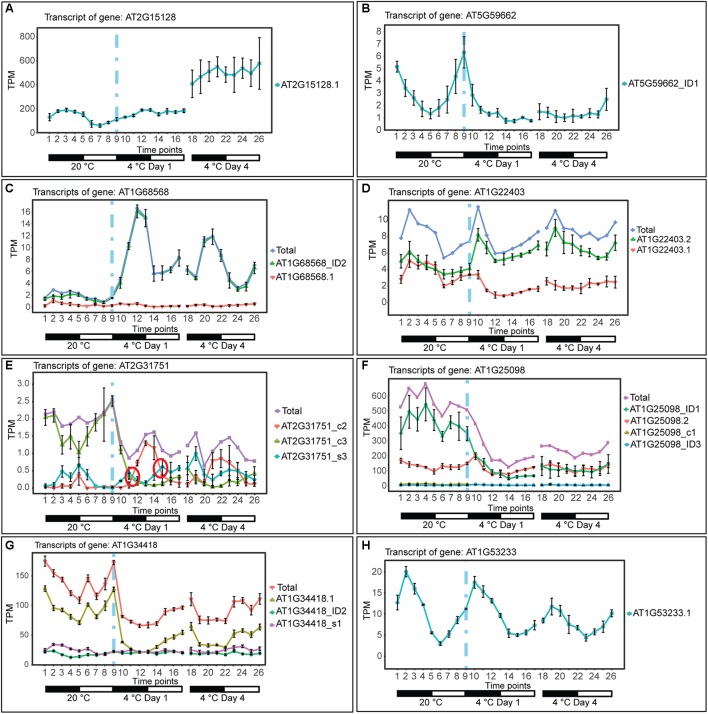
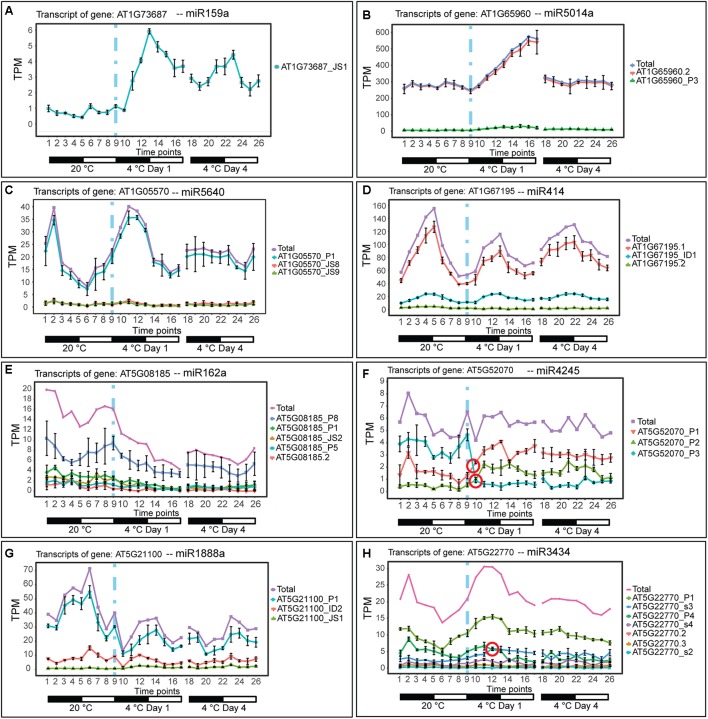
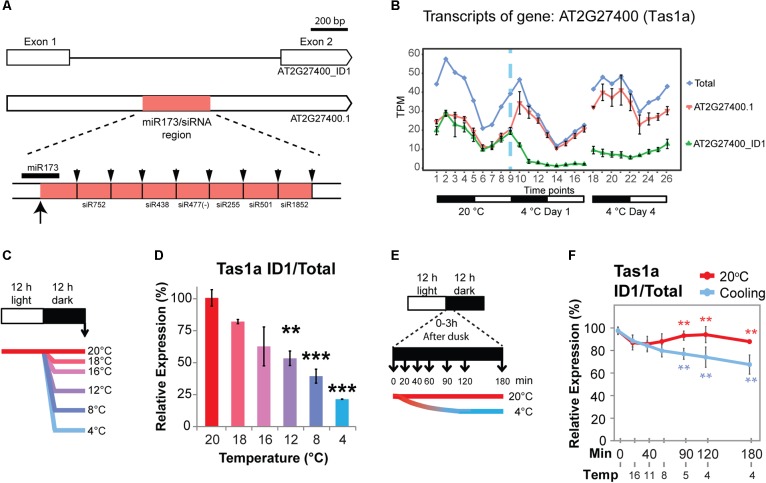
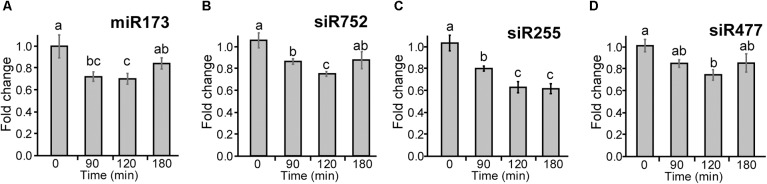
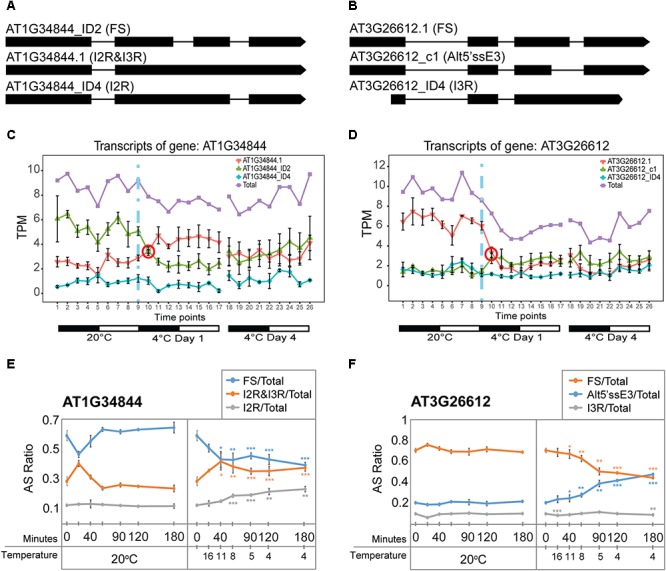
Plants re-program their gene expression when responding to changing environmental conditions. Besides differential gene expression, extensive alternative splicing (AS) of pre-mRNAs and changes in expression of long non-coding RNAs (lncRNAs) are associated with stress responses. RNA-sequencing of a diel time-series of the initial response of Arabidopsis thaliana rosettes to low temperature showed massive and rapid waves of both transcriptional and AS activity in protein-coding genes. We exploited the high diversity of transcript isoforms in AtRTD2 to examine regulation and post-transcriptional regulation of lncRNA gene expression in response to cold stress. We identified 135 lncRNA genes with cold-dependent differential expression (DE) and/or differential alternative splicing (DAS) of lncRNAs including natural antisense RNAs, sORF lncRNAs, and precursors of microRNAs (miRNAs) and trans-acting small-interfering RNAs (tasiRNAs). The high resolution (HR) of the time-series allowed the dynamics of changes in transcription and AS to be determined and identified early and adaptive transcriptional and AS changes in the cold response. Some lncRNA genes were regulated only at the level of AS and using plants grown at different temperatures and a HR time-course of the first 3 h of temperature reduction, we demonstrated that the AS of some lncRNAs is highly sensitive to small temperature changes suggesting tight regulation of expression. In particular, a splicing event in TAS1a which removed an intron that contained the miR173 processing and phased siRNAs generation sites was differentially alternatively spliced in response to cold. The cold-induced reduction of the spliced form of TAS1a and of the tasiRNAs suggests that splicing may enhance production of the siRNAs. Our results identify candidate lncRNAs that may contribute to the regulation of expression that determines the physiological processes essential for acclimation and freezing tolerance.
Keywords: alternative splicing; cold transcriptome; diel time-course; high-resolution RNAseq; long non-coding RNA; primary microRNA.
Figures
Similar articles
-
Dynamic changes in the transcriptome landscape of Arabidopsis thaliana in response to cold stress.Front Plant Sci. 2022 Aug 30;13:983460. doi: 10.3389/fpls.2022.983460. eCollection 2022. Front Plant Sci. 2022. PMID: 36110360 Free PMC article.
-
Identification of small RNAs during cold acclimation in Arabidopsis thaliana.BMC Plant Biol. 2020 Jun 29;20(1):298. doi: 10.1186/s12870-020-02511-3. BMC Plant Biol. 2020. PMID: 32600430 Free PMC article.
-
Rapid and Dynamic Alternative Splicing Impacts the Arabidopsis Cold Response Transcriptome.Plant Cell. 2018 Jul;30(7):1424-1444. doi: 10.1105/tpc.18.00177. Epub 2018 May 15. Plant Cell. 2018. PMID: 29764987 Free PMC article.
-
Research progress on plant noncoding RNAs in response to low-temperature stress.Plant Signal Behav. 2022 Dec 31;17(1):2004035. doi: 10.1080/15592324.2021.2004035. Epub 2021 Dec 19. Plant Signal Behav. 2022. PMID: 34927551 Free PMC article. Review.
-
[Trans-acting short interfering RNAs].Postepy Biochem. 2006;52(3):253-9. Postepy Biochem. 2006. PMID: 17201060 Review. Polish.
Cited by
-
Maize transcriptome profiling reveals low temperatures affect photosynthesis during the emergence stage.Front Plant Sci. 2025 Jan 28;16:1527447. doi: 10.3389/fpls.2025.1527447. eCollection 2025. Front Plant Sci. 2025. PMID: 39935955 Free PMC article.
-
Current perspectives of lncRNAs in abiotic and biotic stress tolerance in plants.Front Plant Sci. 2024 Jan 8;14:1334620. doi: 10.3389/fpls.2023.1334620. eCollection 2023. Front Plant Sci. 2024. PMID: 38259924 Free PMC article. Review.
-
Dynamic changes in the transcriptome landscape of Arabidopsis thaliana in response to cold stress.Front Plant Sci. 2022 Aug 30;13:983460. doi: 10.3389/fpls.2022.983460. eCollection 2022. Front Plant Sci. 2022. PMID: 36110360 Free PMC article.
-
Exosome-Derived LncRNAs in Lung Cancer.Front Oncol. 2020 Sep 23;10:1728. doi: 10.3389/fonc.2020.01728. eCollection 2020. Front Oncol. 2020. PMID: 33072553 Free PMC article. Review.
-
Transcriptomic Analysis Reveals LncRNAs Associated with Flowering of Angelica sinensis during Vernalization.Curr Issues Mol Biol. 2022 Apr 26;44(5):1867-1888. doi: 10.3390/cimb44050128. Curr Issues Mol Biol. 2022. PMID: 35678657 Free PMC article.
References
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials