

Imaging mechanical properties of sub-micron ECM in live zebrafish using Brillouin microscopy
- PMID: 30891356
- PMCID: PMC6420298
- DOI: 10.1364/BOE.10.001420
Imaging mechanical properties of sub-micron ECM in live zebrafish using Brillouin microscopy
Abstract
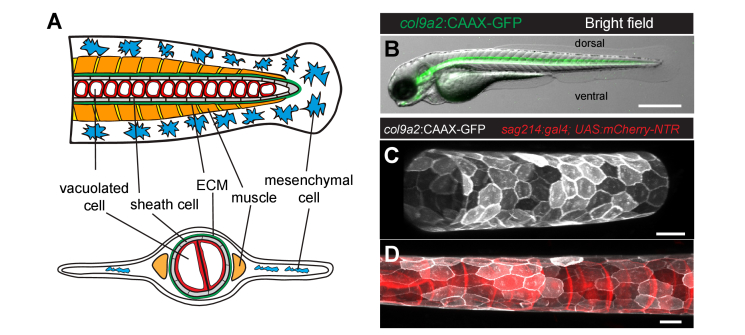
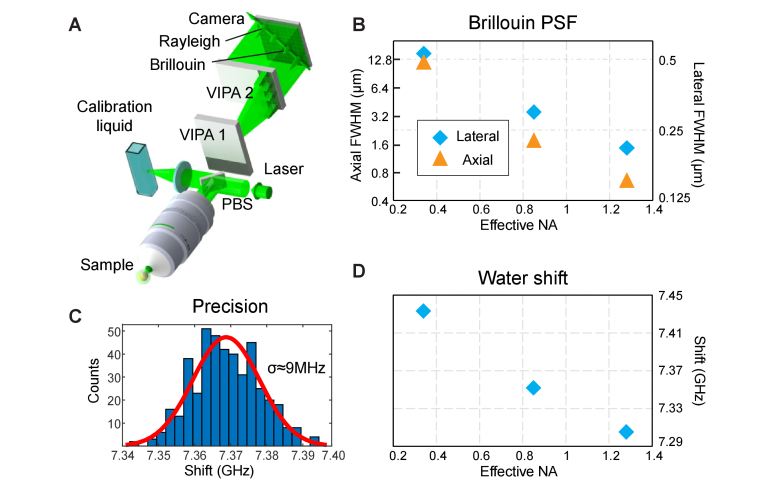
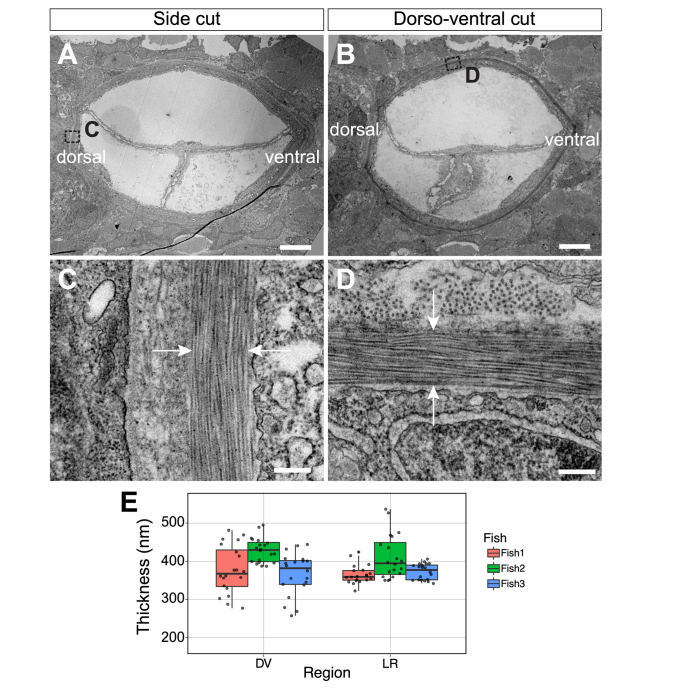
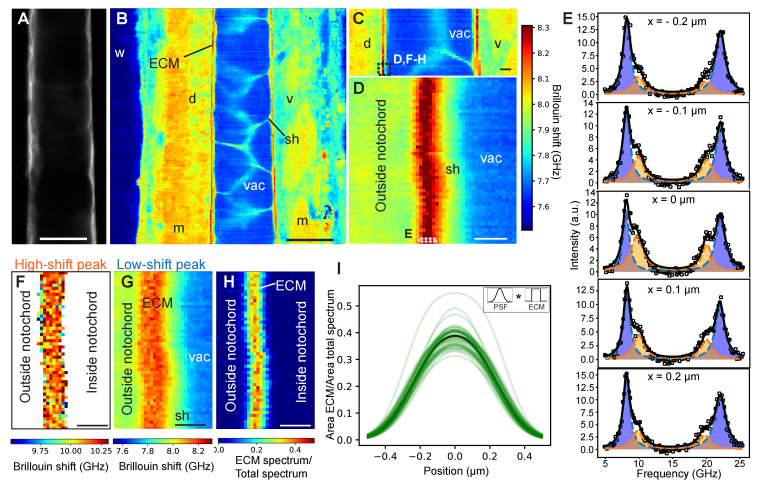
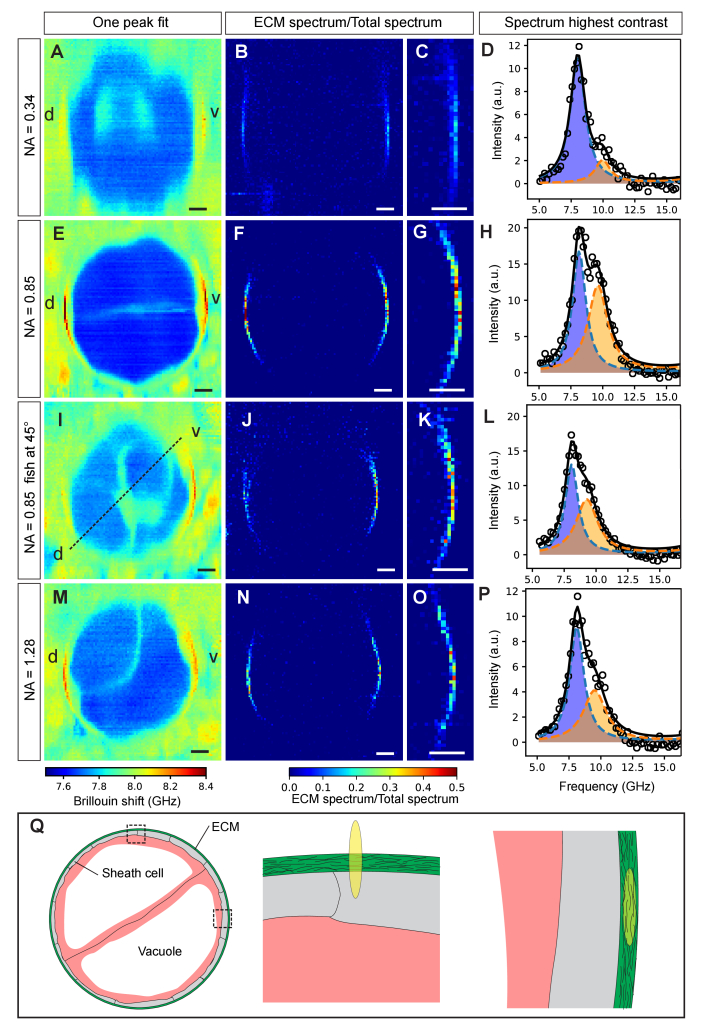
In this work, we quantify the mechanical properties of the extra-cellular matrix (ECM) in live zebrafish using Brillouin microscopy. Optimization of the imaging conditions and parameters, combined with careful spectral analysis, allows us to resolve the thin ECM and distinguish its Brillouin frequency shift, a proxy for mechanical properties, from the surrounding tissue. High-resolution mechanical mapping further enables the direct measurement of the thickness of the ECM label-free and in-vivo. We find the ECM to be ~500 nm thick, and in very good agreement with electron microscopy quantification. Our results open the door for future studies that aim to investigate the role of ECM mechanics for zebrafish morphogenesis and axis elongation.
Conflict of interest statement
The authors declare that there are no conflicts of interest related to this article.
Figures
References
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials