

Electrophysiological impact of thiocyanate on isolated mouse retinal pigment epithelial cells
- PMID: 30892935
- PMCID: PMC6620578
- DOI: 10.1152/ajpcell.00010.2019
Electrophysiological impact of thiocyanate on isolated mouse retinal pigment epithelial cells
Abstract
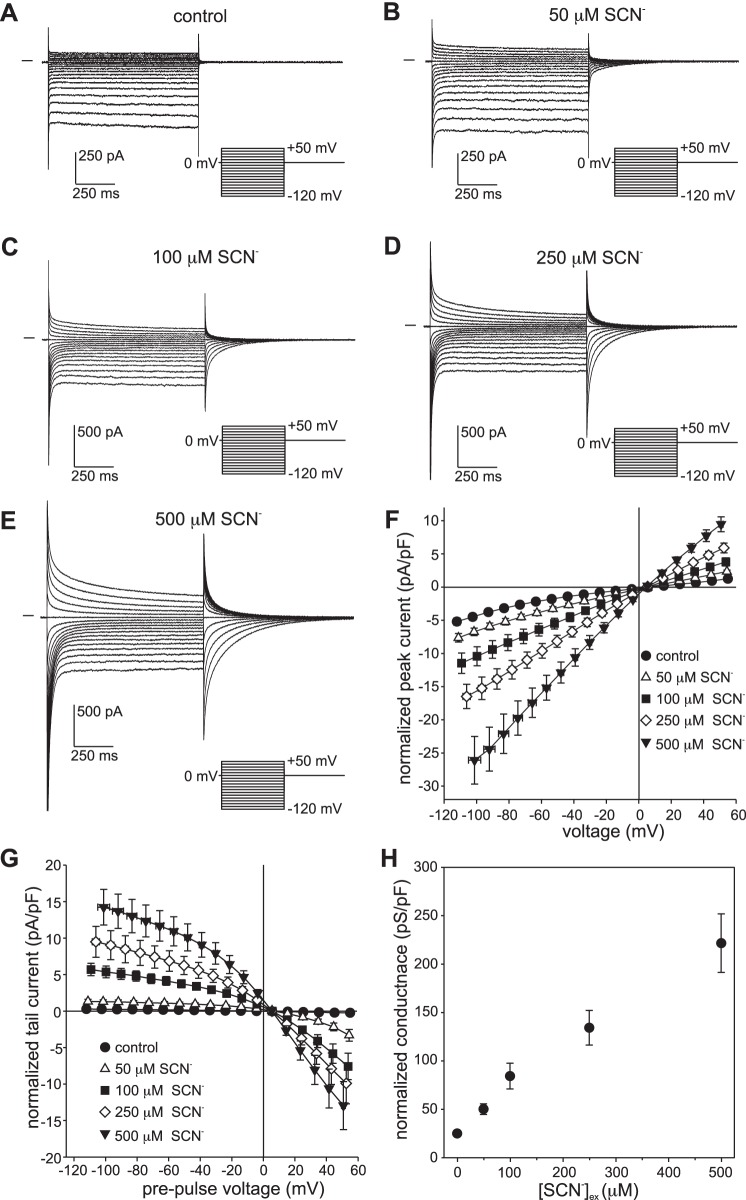
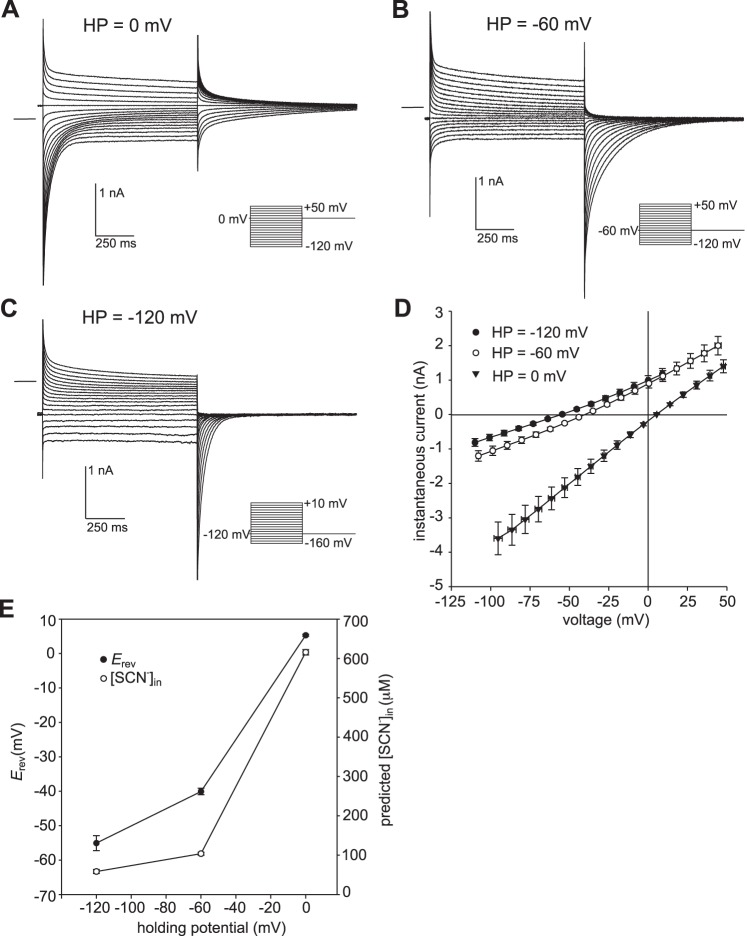
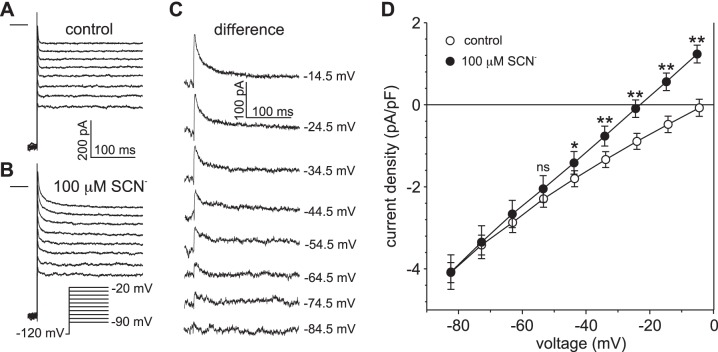
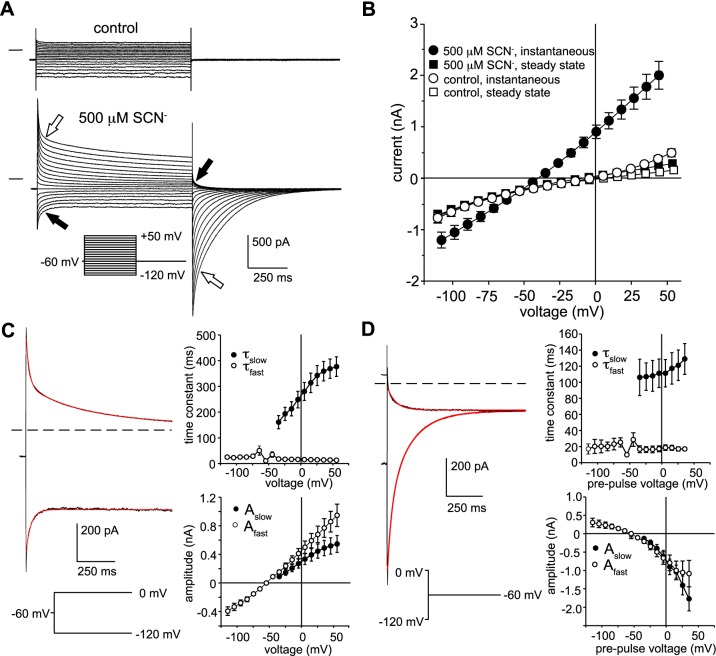
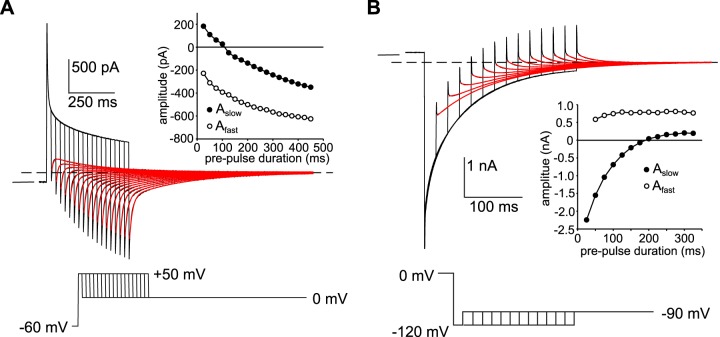
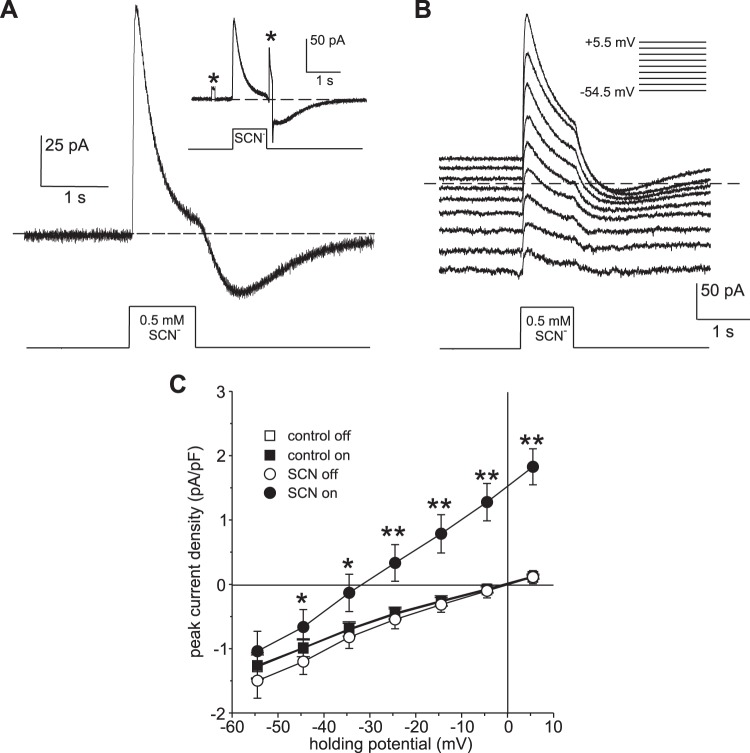
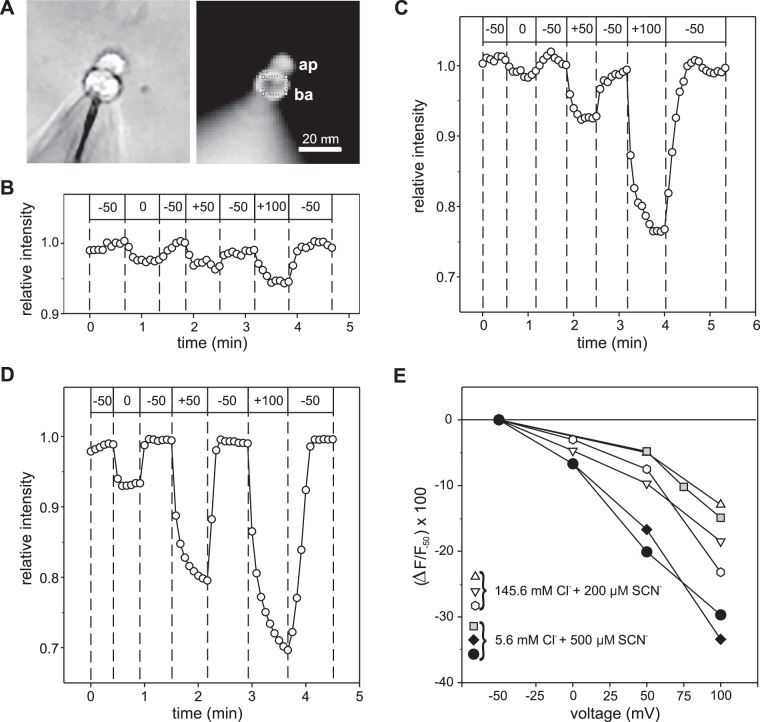
Our recent electrophysiological analysis of mouse retinal pigment epithelial (RPE) cells revealed that in the presence of 10 mM external thiocyanate (SCN-), voltage steps generated large transient currents whose time-dependent decay most likely results from the accumulation or depletion of SCN- intracellularly. In the present study, we investigated the effects of more physiologically relevant concentrations of this biologically active anion. In whole cell recordings of C57BL/6J mouse RPE cells, we found that, over the range of 50 to 500 µM SCN-, the amplitude of transient currents evoked by voltage steps was proportional to the extracellular SCN- concentration. Transient currents were also produced in RPE cells when the membrane potential was held constant and the external SCN- concentration was rapidly increased by pressure-ejecting 500 µM SCN- from a second pipette. Other results indicate that the time dependence of currents produced by both approaches results from a change in driving force due to intracellular SCN- accumulation or depletion. Finally, by applying fluorescence imaging and voltage-clamping techniques to BALB/c mouse RPE cells loaded with the anion-sensitive dye MQAE, we demonstrated that in the presence of 200 or 500 µM extracellular SCN-, depolarizing voltage steps increased the cytoplasmic SCN- concentration to an elevated steady state within several seconds. Collectively, these results indicate that, in the presence of physiological concentrations of SCN- outside the RPE, the conductance and permeability of the RPE cell membranes for SCN- are sufficiently large that SCN- rapidly approaches electrochemical equilibrium within the cytoplasm when the membrane voltage or external SCN- concentration is perturbed.
Keywords: RPE; anion conductance; anion permeability.
Conflict of interest statement
No conflicts of interest, financial or otherwise, are declared by the authors.
Figures
References
-
- Bösl MR, Stein V, Hübner C, Zdebik AA, Jordt SE, Mukhopadhyay AK, Davidoff MS, Holstein AF, Jentsch TJ. Male germ cells and photoreceptors, both dependent on close cell-cell interactions, degenerate upon ClC-2 Cl− channel disruption. EMBO J 20: 1289–1299, 2001. doi: 10.1093/emboj/20.6.1289. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources