Role of the L-PGDS-PGD2-DP1 receptor axis in sleep regulation and neurologic outcomes
- PMID: 30893431
- PMCID: PMC6559173
- DOI: 10.1093/sleep/zsz073
Role of the L-PGDS-PGD2-DP1 receptor axis in sleep regulation and neurologic outcomes
Abstract
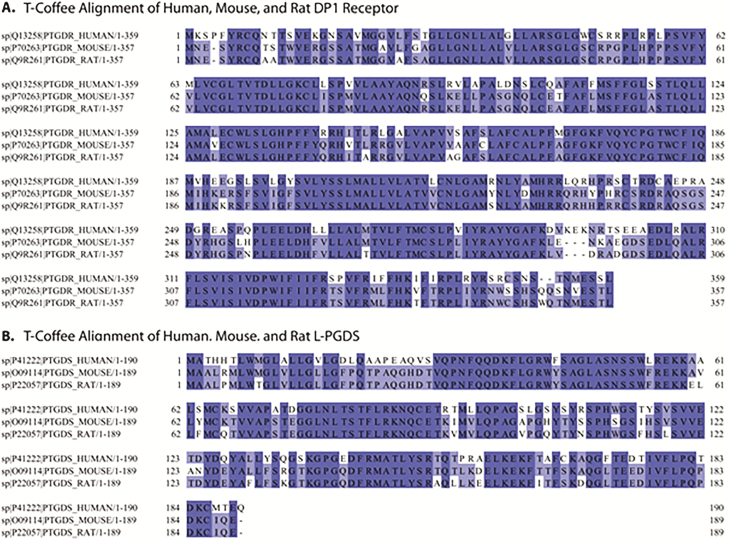
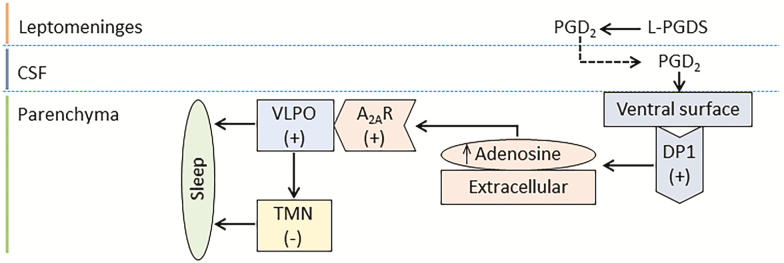
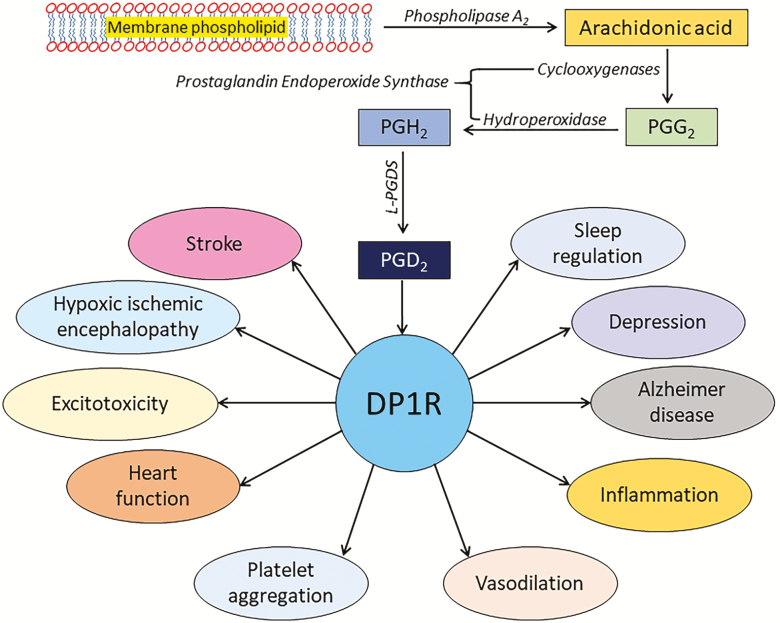
To meet the new challenges of modern lifestyles, we often compromise a good night's sleep. In preclinical models as well as in humans, a chronic lack of sleep is reported to be among the leading causes of various physiologic, psychologic, and neurocognitive deficits. Thus far, various endogenous mediators have been implicated in inter-regulatory networks that collectively influence the sleep-wake cycle. One such mediator is the lipocalin-type prostaglandin D2 synthase (L-PGDS)-Prostaglandin D2 (PGD2)-DP1 receptor (L-PGDS-PGD2-DP1R) axis. Findings in preclinical models confirm that DP1R are predominantly expressed in the sleep-regulating centers. This finding led to the discovery that the L-PGDS-PGD2-DP1R axis is involved in sleep regulation. Furthermore, we showed that the L-PGDS-PGD2-DP1R axis is beneficial in protecting the brain from ischemic stroke. Protein sequence homology was also performed, and it was found that L-PGDS and DP1R share a high degree of homology between humans and rodents. Based on the preclinical and clinical data thus far pertaining to the role of the L-PGDS-PGD2-DP1R axis in sleep regulation and neurologic conditions, there is optimism that this axis may have a high translational potential in human therapeutics. Therefore, here the focus is to review the regulation of the homeostatic component of the sleep process with a special focus on the L-PGDS-PGD2-DP1R axis and the consequences of sleep deprivation on health outcomes. Furthermore, we discuss whether the pharmacological regulation of this axis could represent a tool to prevent sleep disturbances and potentially improve outcomes, especially in patients with acute brain injuries.
Keywords: DP1 receptor; animal models; brain injuries; eicosanoids; inflammation; outcome assessments; poststroke sleep disturbance; prostaglandin D2; prostaglandin receptors; sleep apnea.
© Sleep Research Society 2019. Published by Oxford University Press on behalf of the Sleep Research Society. All rights reserved. For permissions, please e-mail journals.permissions@oup.com.
Figures