

4E-BP1 Is a Tumor Suppressor Protein Reactivated by mTOR Inhibition in Head and Neck Cancer
- PMID: 30894372
- PMCID: PMC6445709
- DOI: 10.1158/0008-5472.CAN-18-1220
4E-BP1 Is a Tumor Suppressor Protein Reactivated by mTOR Inhibition in Head and Neck Cancer
Abstract
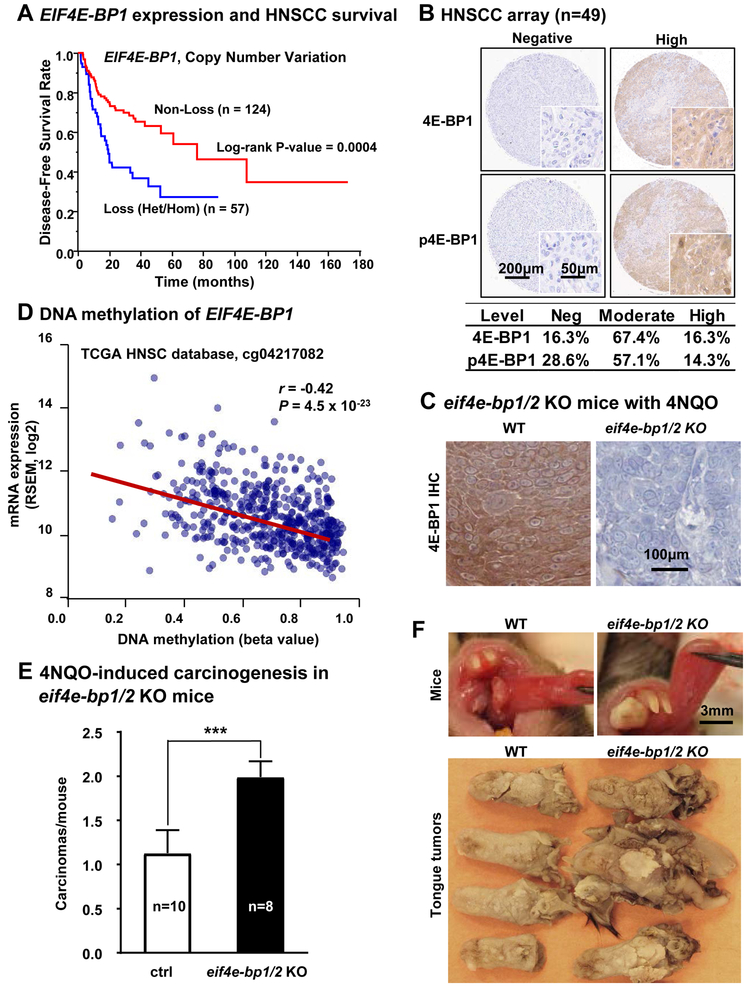
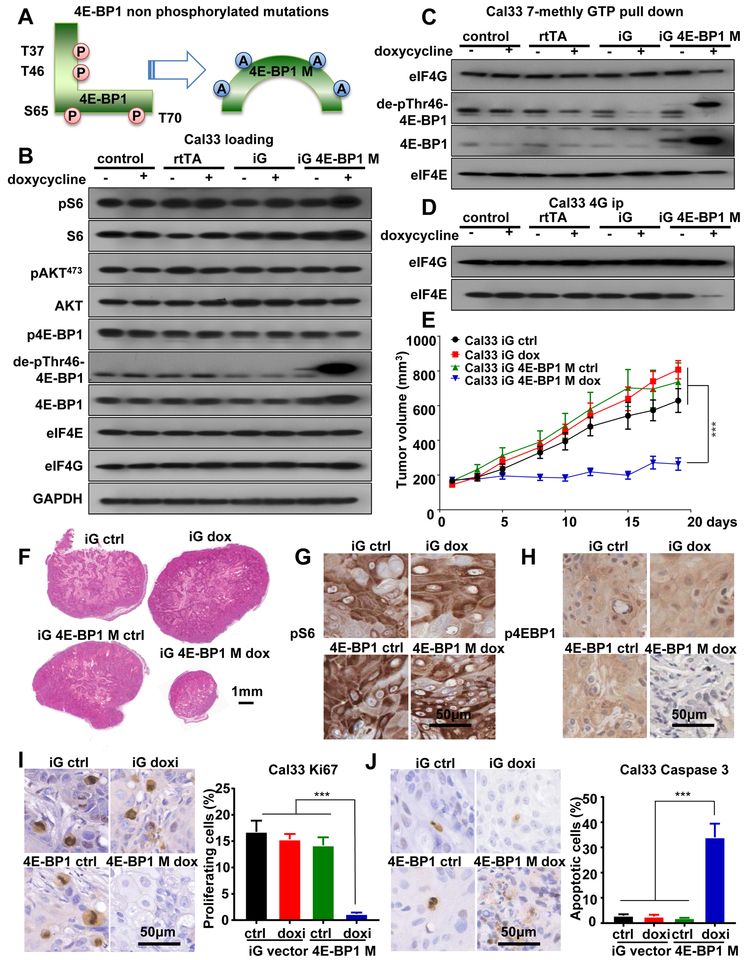
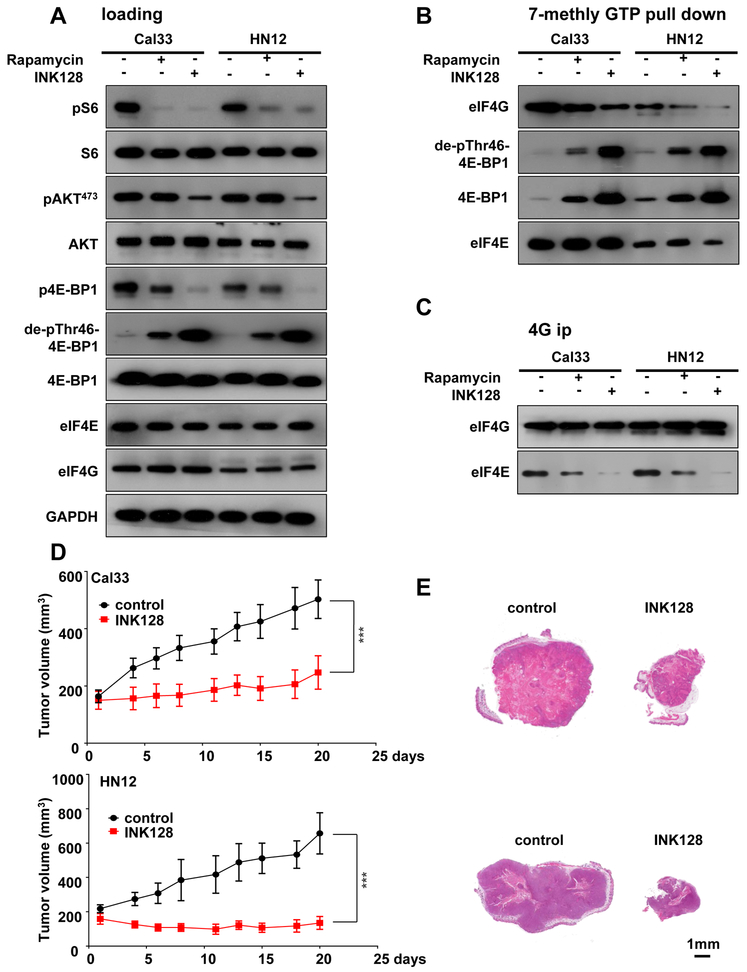
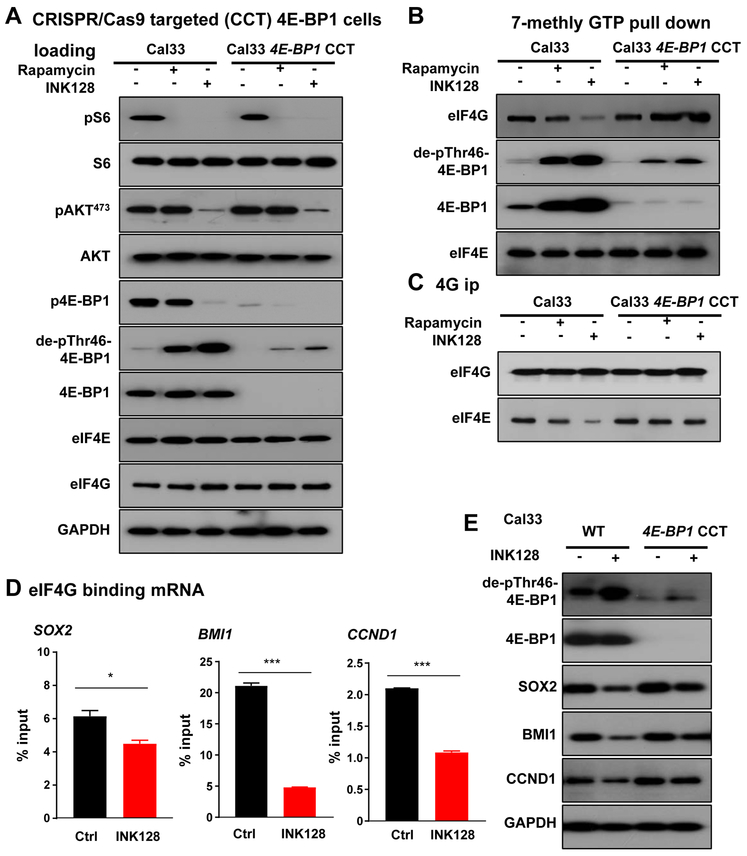
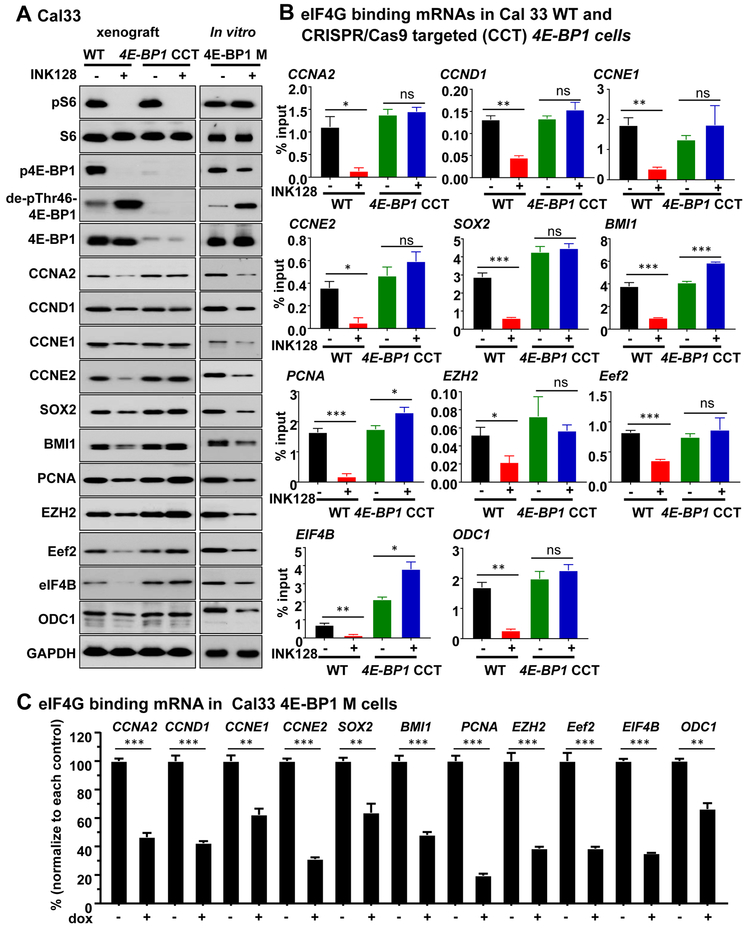
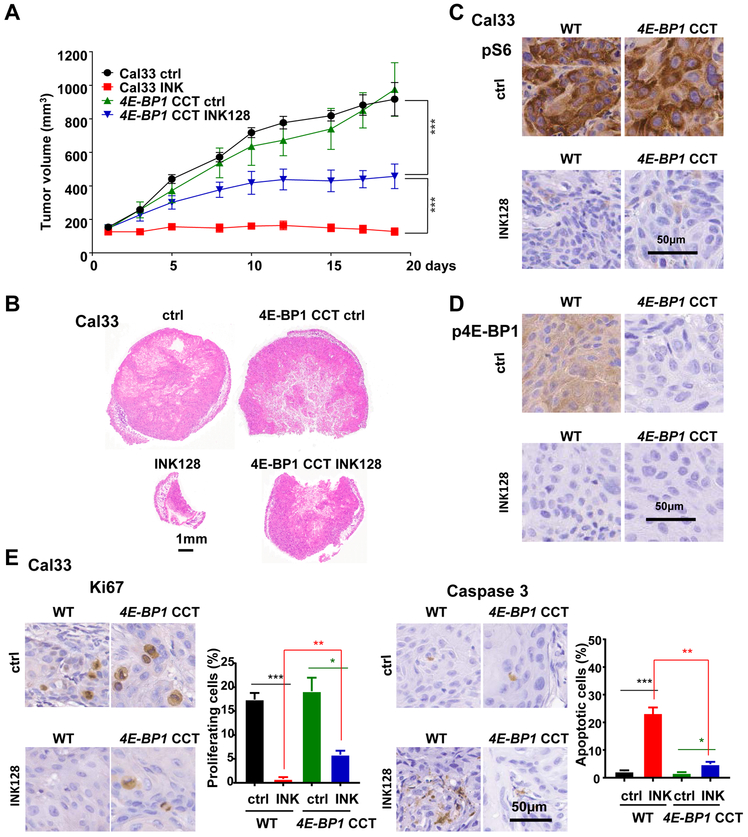
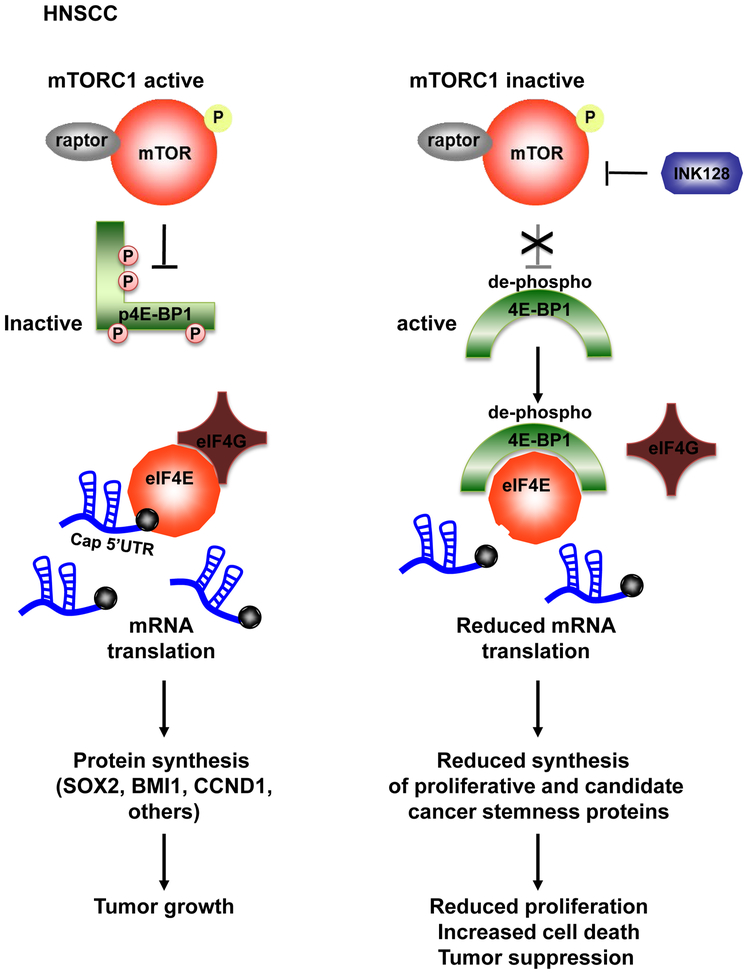
Aberrant activation of the PI3K-mTOR signaling pathway occurs in >80% of head and neck squamous cell carcinomas (HNSCC), and overreliance on this signaling circuit may in turn represent a cancer-specific vulnerability that can be exploited therapeutically. mTOR inhibitors (mTORi) promote tumor regression in genetically defined and chemically induced HNSCC animal models, and encouraging results have been recently reported. However, the mTOR-regulated targets contributing to the clinical response have not yet been identified. Here, we focused on EIF4E-BP1 (4E-BP1), a direct target of mTOR that serves as key effector for protein synthesis. A systematic analysis of genomic alterations in the PIK3CA-mTOR pathway in HNSCC revealed that 4E-BP1 is rarely mutated, but at least one 4E-BP1 gene copy is lost in over 35% of the patients with HNSCC, correlating with decreased 4E-BP1 protein expression. 4E-BP1 gene copy number loss correlated with poor disease-free and overall survival. Aligned with a tumor-suppressive role, 4e-bp1/2 knockout mice formed larger and more lesions in models of HNSCC carcinogenesis. mTORi treatment or conditional expression of a mutant 4E-BP1 that cannot be phosphorylated by mTOR was sufficient to disrupt the translation-initiation complex and prevent tumor growth. Furthermore, CRISPR/Cas9-targeted 4E-BP1 HNSCC cells resulted in reduced sensitivity to mTORi in vitro and in vivo. Overall, these findings indicate that in HNSCC, mTOR persistently restrains 4E-BP1 via phosphorylation and that mTORi can restore the tumor-suppressive function of 4E-BP1. Our findings also support 4E-BP1 expression and phosphorylation status as a mechanistic biomarker of mTORi sensitivity in patients with HNSCC. SIGNIFICANCE: These findings suggest that EIF4E-BP1 acts as a tumor suppressor in HNSCC and that 4E-BP1 dephosphorylation mediates the therapeutic response to mTORi, providing a mechanistic biomarker for future precision oncology trials.
©2019 American Association for Cancer Research.
Conflict of interest statement
Competing financial interests
On behalf of all other authors, the corresponding author states that there is no conflict of interest
Figures
References
-
- Siegel RL, Miller KD, Jemal A. Cancer statistics, 2018. CA: a cancer journal for clinicians; 2018;68:7–30 - PubMed
-
- Leemans CR, Braakhuis BJ, Brakenhoff RH. The molecular biology of head and neck cancer. Nat Rev Cancer 2011;11:9–22 - PubMed
-
- Amornphimoltham P, Patel V, Sodhi A, Nikitakis NG, Sauk JJ, Sausville EA, et al. Mammalian target of rapamycin, a molecular target in squamous cell carcinomas of the head and neck. Cancer research 2005;65:9953–61 - PubMed
-
- Molinolo AA, Hewitt SM, Amornphimoltham P, Keelawat S, Rangdaeng S, Meneses Garcia A, et al. Dissecting the Akt/mammalian target of rapamycin signaling network: emerging results from the head and neck cancer tissue array initiative. Clinical cancer research : an official journal of the American Association for Cancer Research 2007;13:4964–73 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
