

Cell Entry-Independent Role for the Reovirus μ1 Protein in Regulating Necroptosis and the Accumulation of Viral Gene Products
- PMID: 30894465
- PMCID: PMC6532083
- DOI: 10.1128/JVI.00199-19
Cell Entry-Independent Role for the Reovirus μ1 Protein in Regulating Necroptosis and the Accumulation of Viral Gene Products
Abstract
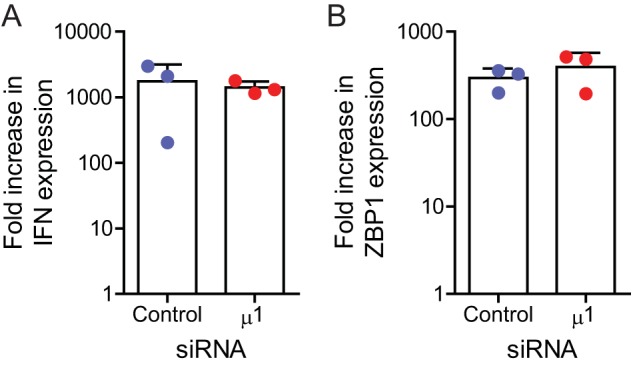
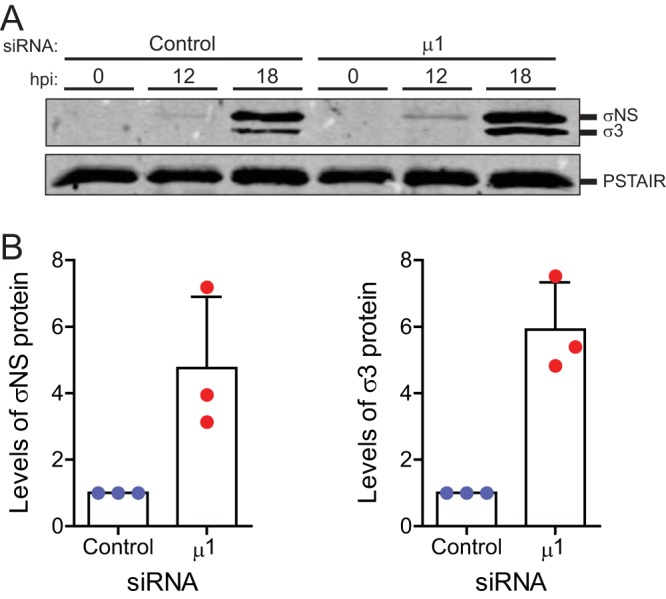
The reovirus outer capsid protein μ1 regulates cell death in infected cells. To distinguish between the roles of incoming, capsid-associated, and newly synthesized μ1, we used small interfering RNA (siRNA)-mediated knockdown. Loss of newly synthesized μ1 protein does not affect apoptotic cell death in HeLa cells but enhances necroptosis in L929 cells. Knockdown of μ1 also affects aspects of viral replication. We found that, while μ1 knockdown results in diminished release of infectious viral progeny from infected cells, viral minus-strand RNA, plus-strand RNA, and proteins that are not targeted by the μ1 siRNA accumulate to a greater extent than in control siRNA-treated cells. Furthermore, we observed a decrease in sensitivity of these viral products to inhibition by guanidine hydrochloride (GuHCl) (which targets minus-strand synthesis to produce double-stranded RNA) when μ1 is knocked down. Following μ1 knockdown, cell death is also less sensitive to treatment with GuHCl. Our studies suggest that the absence of μ1 allows enhanced transcriptional activity of newly synthesized cores and the consequent accumulation of viral gene products. We speculate that enhanced accumulation and detection of these gene products due to μ1 knockdown potentiates receptor-interacting protein 3 (RIP3)-dependent cell death.IMPORTANCE We used mammalian reovirus as a model to study how virus infections result in cell death. Here, we sought to determine how viral factors regulate cell death. Our work highlights a previously unknown role for the reovirus outer capsid protein μ1 in limiting the induction of a necrotic form of cell death called necroptosis. Induction of cell death by necroptosis requires the detection of viral gene products late in infection; μ1 limits cell death by this mechanism because it prevents excessive accumulation of viral gene products that trigger cell death.
Keywords: cell death; reovirus.
Copyright © 2019 American Society for Microbiology.
Figures








References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
