

Fibroblast growth factor 21 controls mitophagy and muscle mass
- PMID: 30895728
- PMCID: PMC6596457
- DOI: 10.1002/jcsm.12409
Fibroblast growth factor 21 controls mitophagy and muscle mass
Abstract
Background: Skeletal muscle is a plastic tissue that adapts to changes in exercise, nutrition, and stress by secreting myokines and myometabolites. These muscle-secreted factors have autocrine, paracrine, and endocrine effects, contributing to whole body homeostasis. Muscle dysfunction in aging sarcopenia, cancer cachexia, and diabetes is tightly correlated with the disruption of the physiological homeostasis at the whole body level. The expression levels of the myokine fibroblast growth factor 21 (FGF21) are very low in normal healthy muscles. However, fasting, ER stress, mitochondrial myopathies, and metabolic disorders induce its release from muscles. Although our understanding of the systemic effects of muscle-derived FGF21 is exponentially increasing, the direct contribution of FGF21 to muscle function has not been investigated yet.
Methods: Muscle-specific FGF21 knockout mice were generated to investigate the consequences of FGF21 deletion concerning skeletal muscle mass and force. To identify the mechanisms underlying FGF21-dependent adaptations in skeletal muscle during starvation, the study was performed on muscles collected from both fed and fasted adult mice. In vivo overexpression of FGF21 was performed in skeletal muscle to assess whether FGF21 is sufficient per se to induce muscle atrophy.
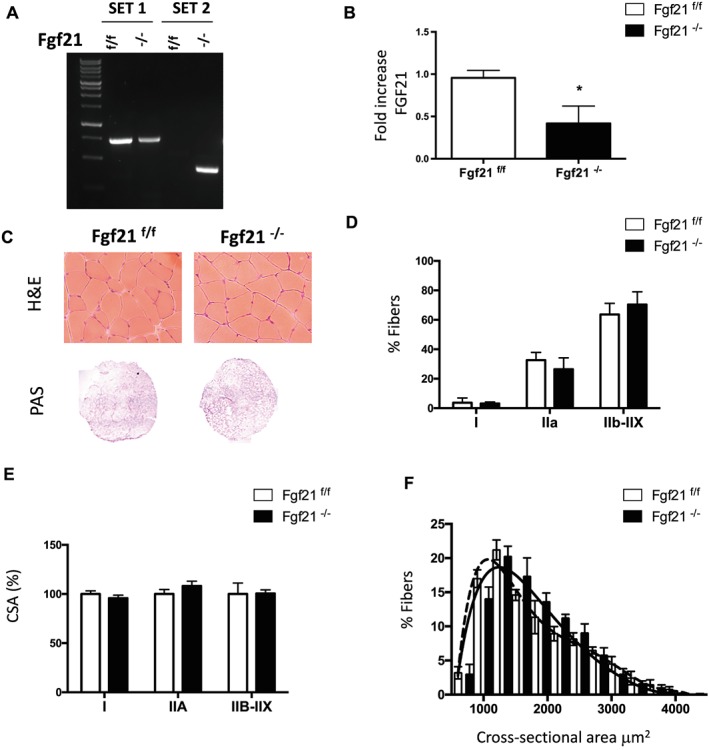
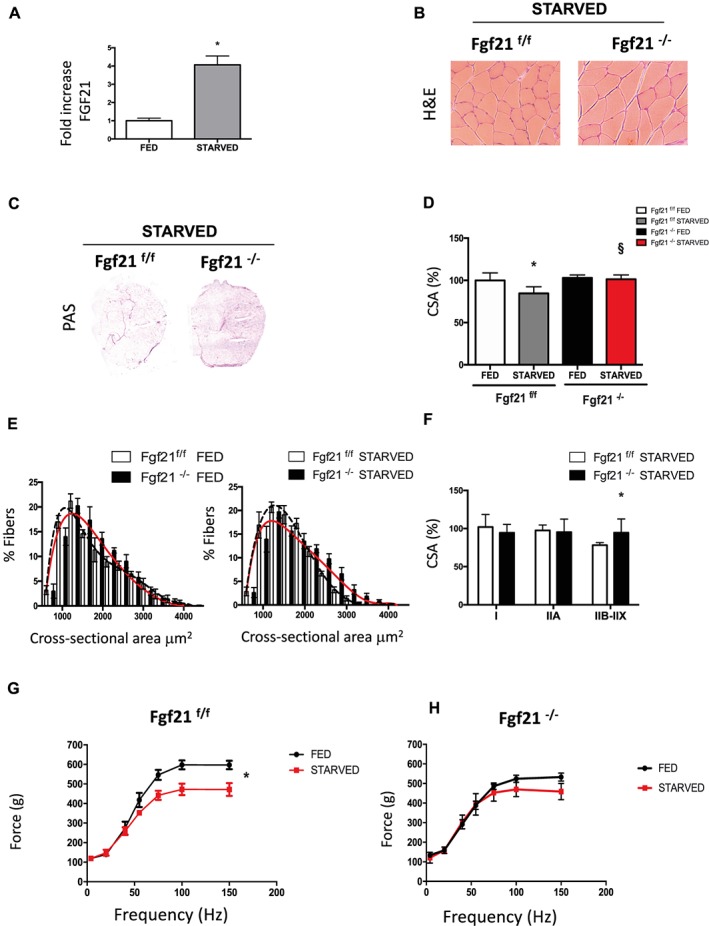
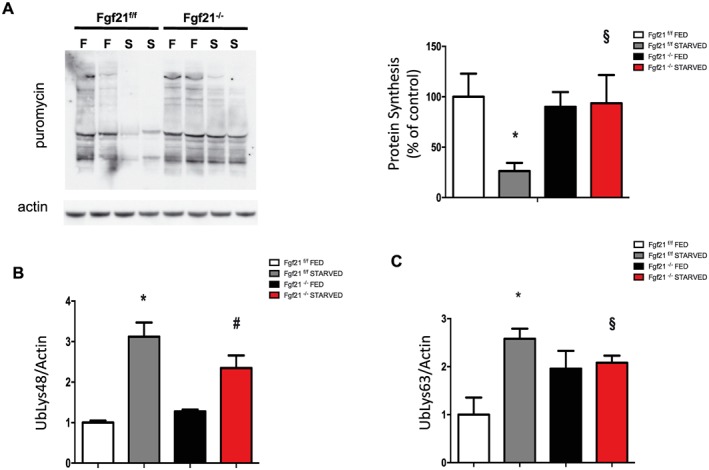
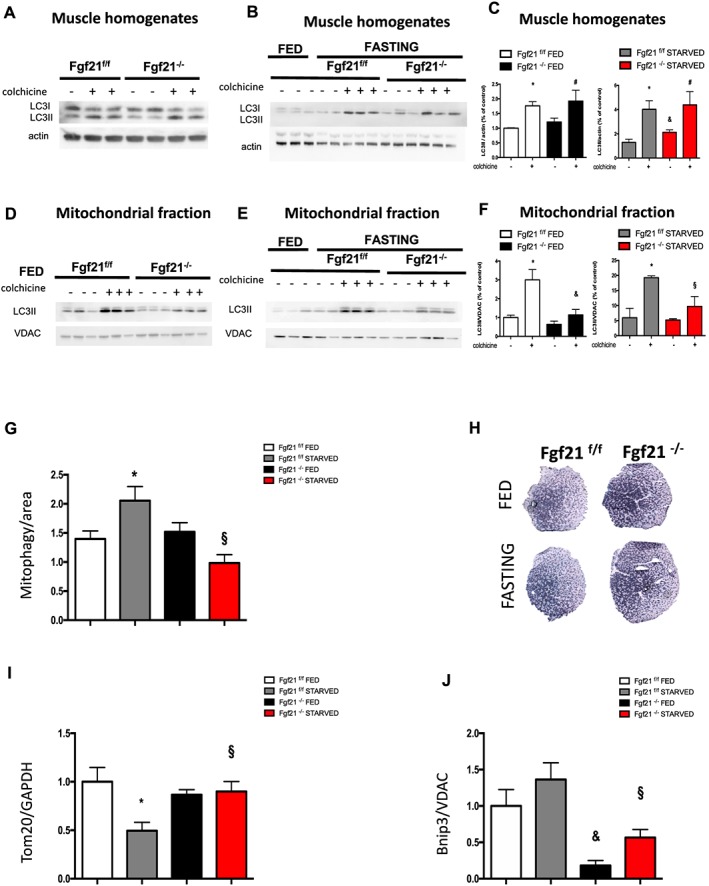
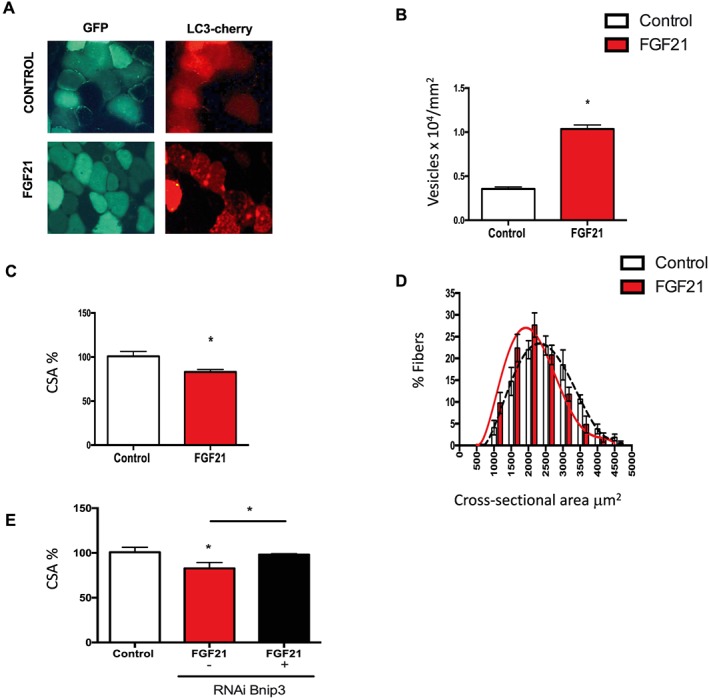
Results: We show that FGF21 does not contribute to muscle homeostasis in basal conditions in terms of fibre type distribution, fibre size, and muscle force. In contrast, FGF21 is required for fasting-induced muscle atrophy and weakness. The mass of isolated muscles from control-fasted mice was reduced by 15-25% (P < 0.05) compared with fed control mice. FGF21-null muscles, however, were significantly protected from muscle loss and weakness during fasting. Such important protection is due to the maintenance of protein synthesis rate in knockout muscles during fasting compared with a 70% reduction in control-fasted muscles (P < 0.01), together with a significant reduction of the mitophagy flux via the regulation of the mitochondrial protein Bnip3. The contribution of FGF21 to the atrophy programme was supported by in vivo FGF21 overexpression in muscles, which was sufficient to induce autophagy and muscle loss by 15% (P < 0.05). Bnip3 inhibition protected against FGF21-dependent muscle wasting in adult animals (P < 0.05).
Conclusions: FGF21 is a novel player in the regulation of muscle mass that requires the mitophagy protein Bnip3.
Keywords: Autophagy; Bnip3; FGF21; Mitophagy; Muscle atrophy; Myokine.
© 2019 The Authors. Journal of Cachexia, Sarcopenia and Muscle published by John Wiley & Sons Ltd on behalf of the Society on Sarcopenia, Cachexia and Wasting Disorders.
Conflict of interest statement
None declared.
Figures
Comment in
-
Comment on: "Fibroblast growth factor 21 controls mitophagy and muscle mass" by Oost et al.J Cachexia Sarcopenia Muscle. 2020 Feb;11(1):336-337. doi: 10.1002/jcsm.12465. Epub 2019 Jul 15. J Cachexia Sarcopenia Muscle. 2020. PMID: 31307122 Free PMC article. No abstract available.
-
The authors reply: Letter on: "Fibroblast growth factor 21 controls mitophagy and muscle mass" by Oost et al.J Cachexia Sarcopenia Muscle. 2020 Feb;11(1):338-340. doi: 10.1002/jcsm.12500. Epub 2019 Nov 6. J Cachexia Sarcopenia Muscle. 2020. PMID: 31692288 Free PMC article. No abstract available.
References
-
- Volaklis KA, Halle M, Meisinger C. Muscular strength as a strong predictor of mortality: a narrative review. Eur J Intern Med 2015;26:303–310. - PubMed
-
- Rai M, Demontis F. Systemic nutrient and stress signaling via myokines and myometabolites. Annu Rev Physiol 2016;78:85–107. - PubMed
-
- Whitham M, Febbraio MA. The ever‐expanding myokinome: discovery challenges and therapeutic implications. Nat Rev Drug Discov 2016;15:719–729. - PubMed
-
- Pedersen BK, Steensberg A, Fischer C, Keller C, Keller P, Plomgaard P, et al. Searching for the exercise factor: is IL‐6 a candidate? J Muscle Res Cell Motil 2003;24:113–119. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
