

Comprehensive profiling of the STE20 kinase family defines features essential for selective substrate targeting and signaling output
- PMID: 30897078
- PMCID: PMC6445471
- DOI: 10.1371/journal.pbio.2006540
Comprehensive profiling of the STE20 kinase family defines features essential for selective substrate targeting and signaling output
Abstract
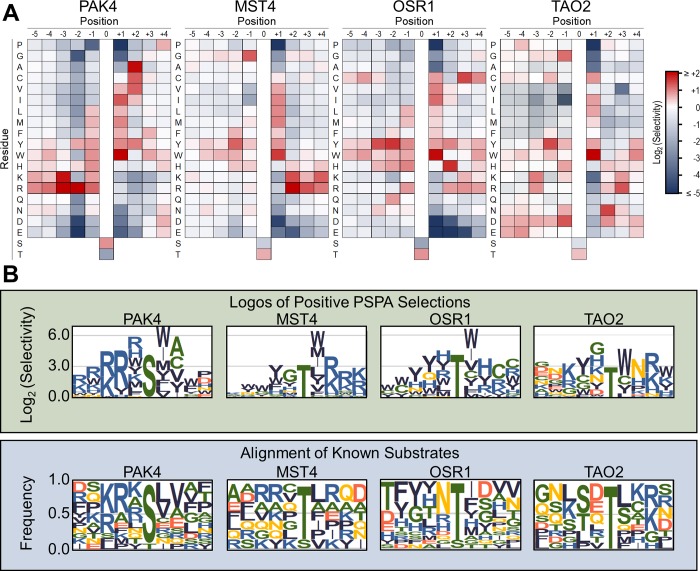
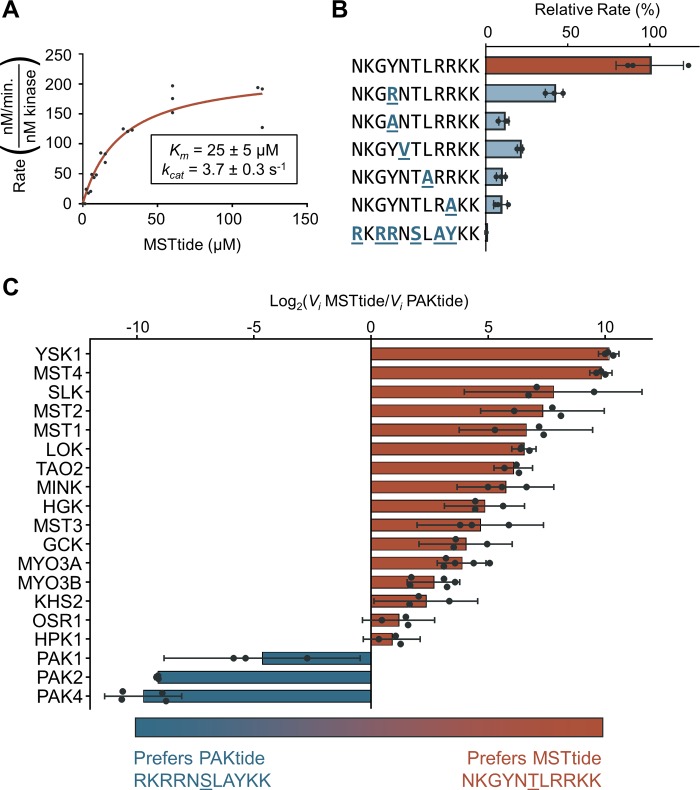
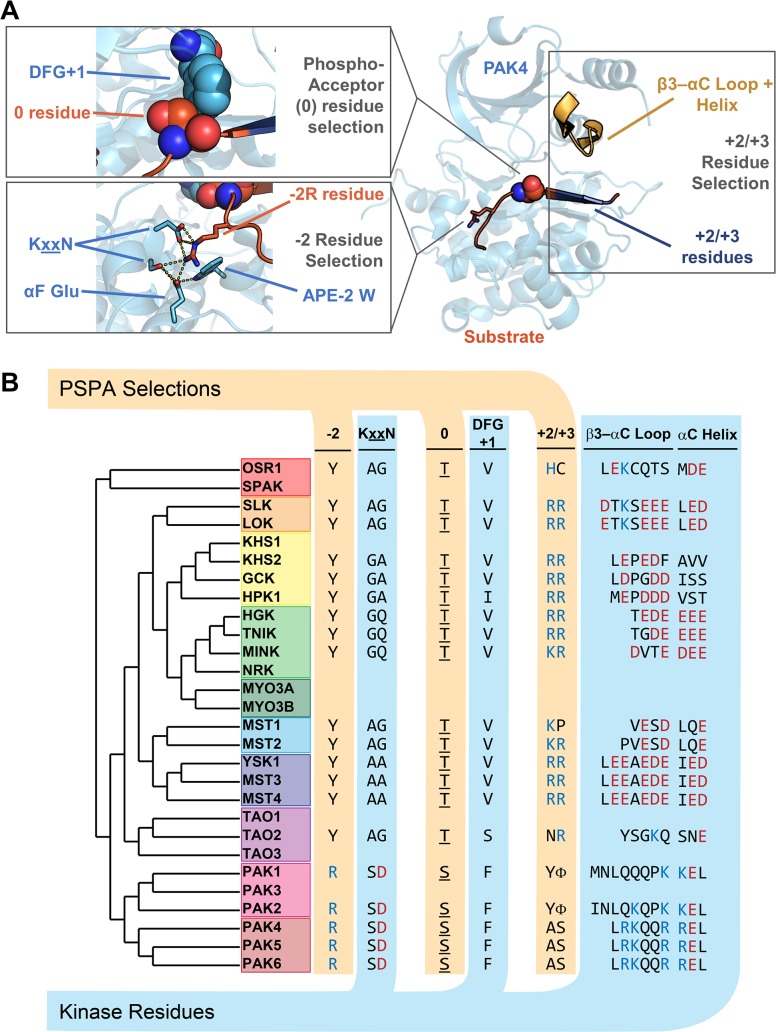
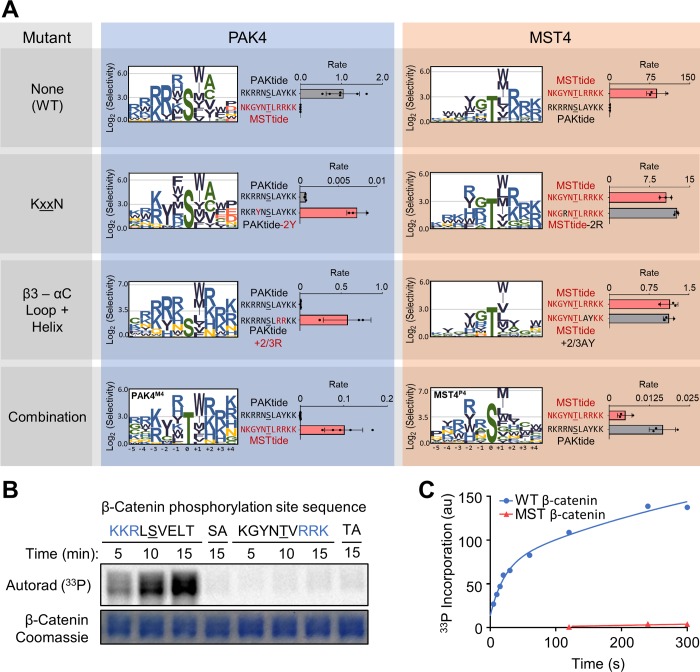
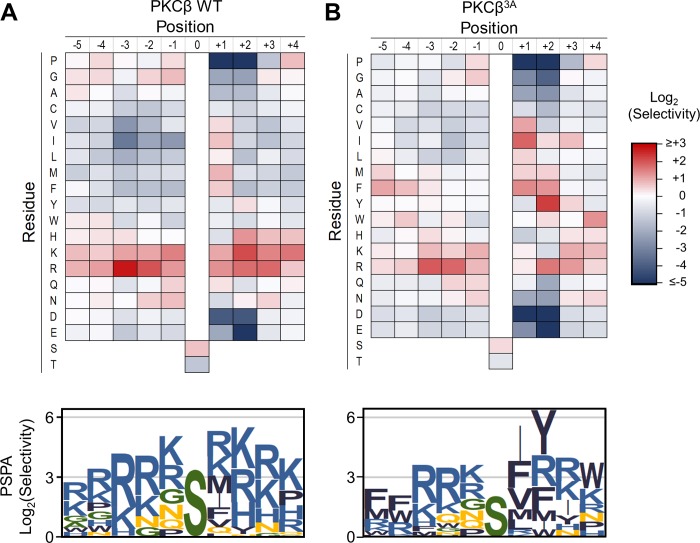
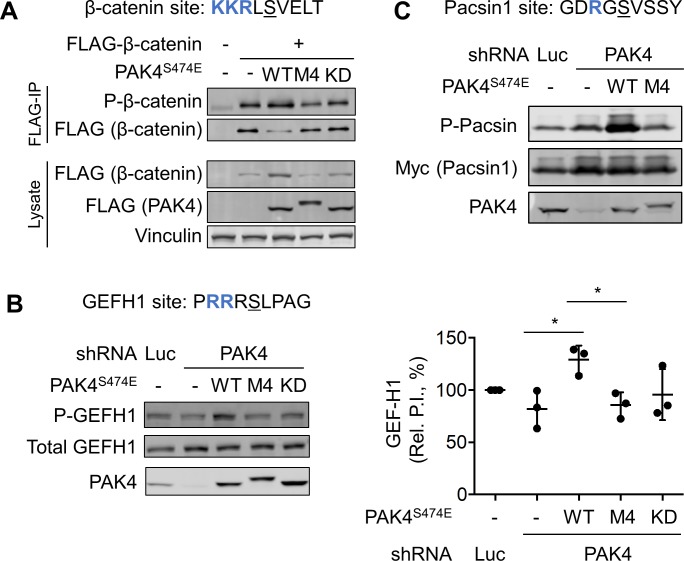
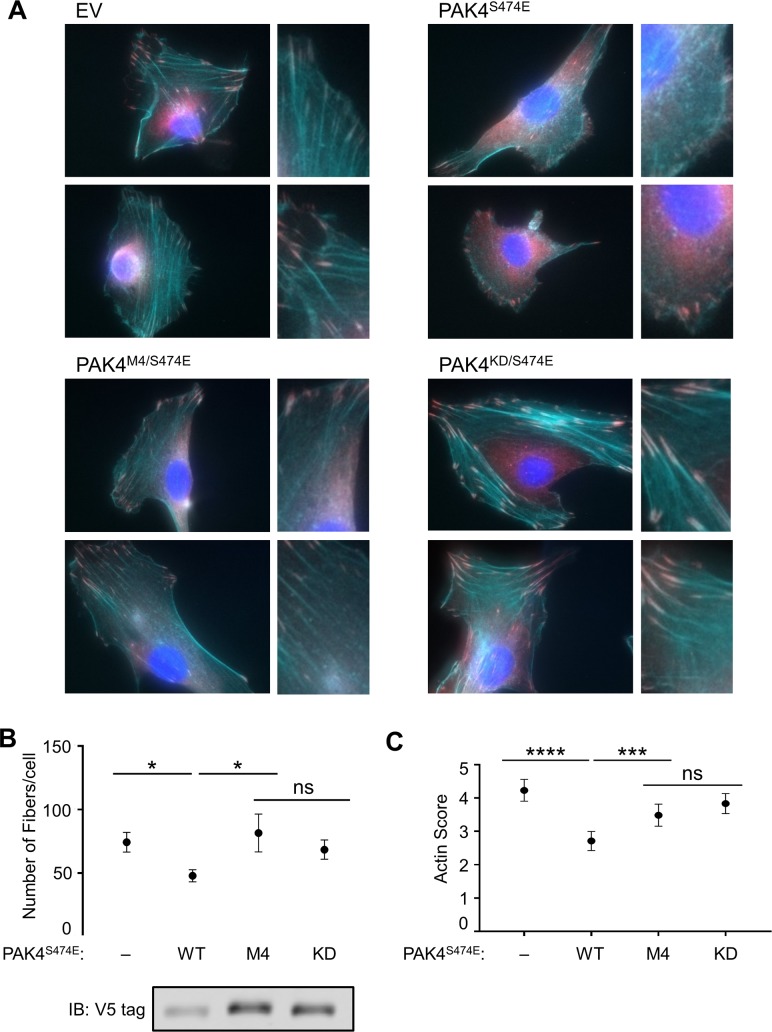
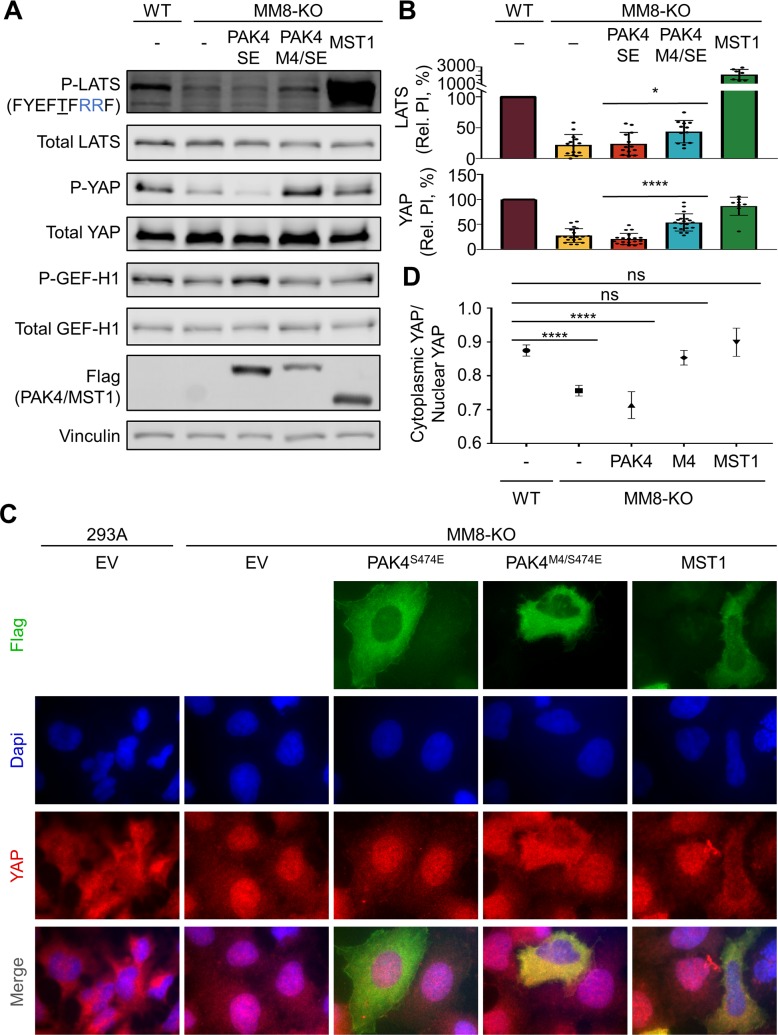
Specificity within protein kinase signaling cascades is determined by direct and indirect interactions between kinases and their substrates. While the impact of localization and recruitment on kinase-substrate targeting can be readily assessed, evaluating the relative importance of direct phosphorylation site interactions remains challenging. In this study, we examine the STE20 family of protein serine-threonine kinases to investigate basic mechanisms of substrate targeting. We used peptide arrays to define the phosphorylation site specificity for the majority of STE20 kinases and categorized them into four distinct groups. Using structure-guided mutagenesis, we identified key specificity-determining residues within the kinase catalytic cleft, including an unappreciated role for the kinase β3-αC loop region in controlling specificity. Exchanging key residues between the STE20 kinases p21-activated kinase 4 (PAK4) and Mammalian sterile 20 kinase 4 (MST4) largely interconverted their phosphorylation site preferences. In cells, a reprogrammed PAK4 mutant, engineered to recognize MST substrates, failed to phosphorylate PAK4 substrates or to mediate remodeling of the actin cytoskeleton. In contrast, this mutant could rescue signaling through the Hippo pathway in cells lacking multiple MST kinases. These observations formally demonstrate the importance of catalytic site specificity for directing protein kinase signal transduction pathways. Our findings further suggest that phosphorylation site specificity is both necessary and sufficient to mediate distinct signaling outputs of STE20 kinases and imply broad applicability to other kinase signaling systems.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Identification of a major determinant for serine-threonine kinase phosphoacceptor specificity.Mol Cell. 2014 Jan 9;53(1):140-7. doi: 10.1016/j.molcel.2013.11.013. Epub 2013 Dec 26. Mol Cell. 2014. PMID: 24374310 Free PMC article.
-
Human Ste20 homologue hPAK1 links GTPases to the JNK MAP kinase pathway.Curr Biol. 1996 May 1;6(5):598-605. doi: 10.1016/s0960-9822(02)00546-8. Curr Biol. 1996. PMID: 8805275
-
Activation loop phosphorylation of ERK3/ERK4 by group I p21-activated kinases (PAKs) defines a novel PAK-ERK3/4-MAPK-activated protein kinase 5 signaling pathway.J Biol Chem. 2011 Feb 25;286(8):6470-8. doi: 10.1074/jbc.M110.181529. Epub 2010 Dec 22. J Biol Chem. 2011. PMID: 21177870 Free PMC article.
-
Signaling, Regulation, and Specificity of the Type II p21-activated Kinases.J Biol Chem. 2015 May 22;290(21):12975-83. doi: 10.1074/jbc.R115.650416. Epub 2015 Apr 8. J Biol Chem. 2015. PMID: 25855792 Free PMC article. Review.
-
Regulation of mammalian Ste20 (Mst) kinases.Trends Biochem Sci. 2015 Mar;40(3):149-56. doi: 10.1016/j.tibs.2015.01.001. Epub 2015 Feb 6. Trends Biochem Sci. 2015. PMID: 25665457 Free PMC article. Review.
Cited by
-
MST4 kinase suppresses gastric tumorigenesis by limiting YAP activation via a non-canonical pathway.J Exp Med. 2020 Jun 1;217(6):e20191817. doi: 10.1084/jem.20191817. J Exp Med. 2020. PMID: 32271880 Free PMC article.
-
Pleiotropic functions of TAO kinases and their dysregulation in neurological disorders.Sci Signal. 2024 Jan 2;17(817):eadg0876. doi: 10.1126/scisignal.adg0876. Epub 2024 Jan 2. Sci Signal. 2024. PMID: 38166033 Free PMC article. Review.
-
Multi-Omics Analysis Revealed That TAOK1 Can Be Used as a Prognostic Marker and Target in a Variety of Tumors, Especially in Cervical Cancer.Onco Targets Ther. 2025 Mar 14;18:335-353. doi: 10.2147/OTT.S506582. eCollection 2025. Onco Targets Ther. 2025. PMID: 40109409 Free PMC article.
-
Recognition of physiological phosphorylation sites by p21-activated kinase 4.J Struct Biol. 2020 Sep 1;211(3):107553. doi: 10.1016/j.jsb.2020.107553. Epub 2020 Jun 23. J Struct Biol. 2020. PMID: 32585314 Free PMC article.
-
Epinephrine inhibits PI3Kα via the Hippo kinases.Cell Rep. 2023 Dec 26;42(12):113535. doi: 10.1016/j.celrep.2023.113535. Epub 2023 Dec 4. Cell Rep. 2023. PMID: 38060450 Free PMC article.
References
-
- Pinna LA, Ruzzene M. How do protein kinases recognize their substrates? Biochim. Biophys. Acta. 1996;1314: 191–225. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
