

The vitamin E derivative garcinoic acid from Garcinia kola nut seeds attenuates the inflammatory response
- PMID: 30897408
- PMCID: PMC6426704
- DOI: 10.1016/j.redox.2019.101166
The vitamin E derivative garcinoic acid from Garcinia kola nut seeds attenuates the inflammatory response
Abstract
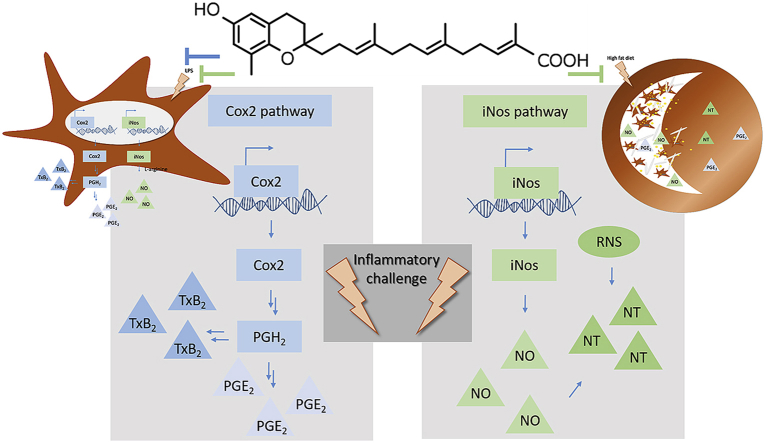
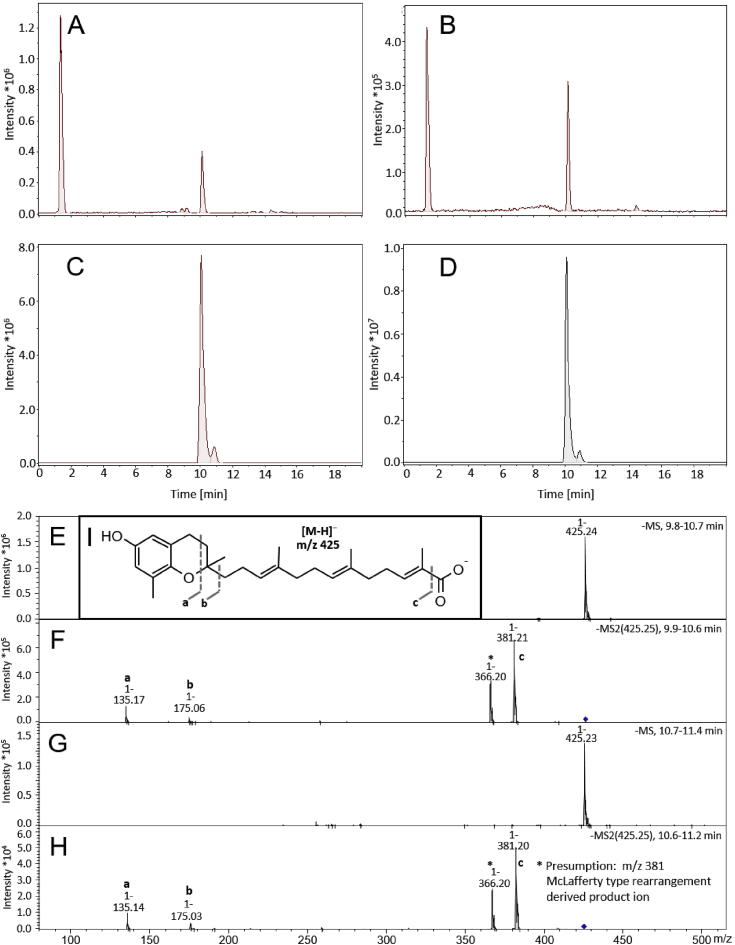
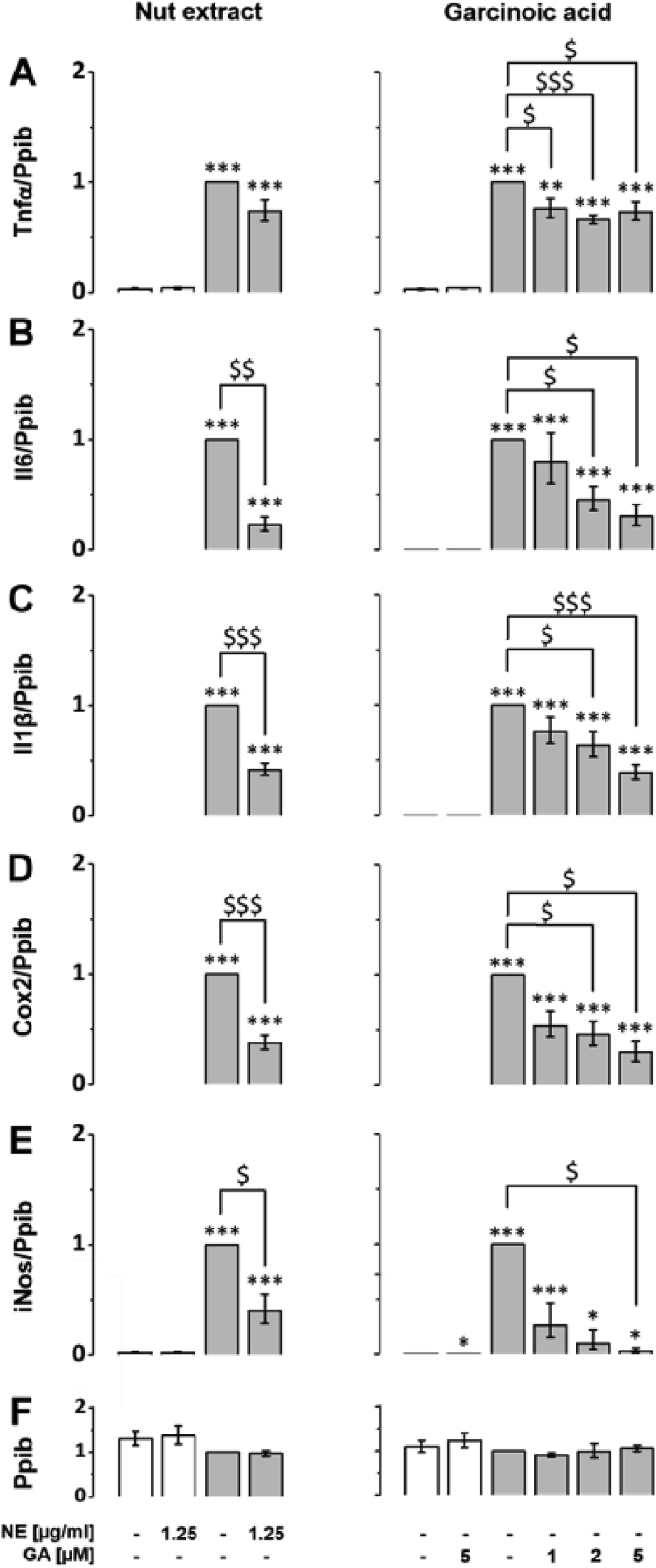
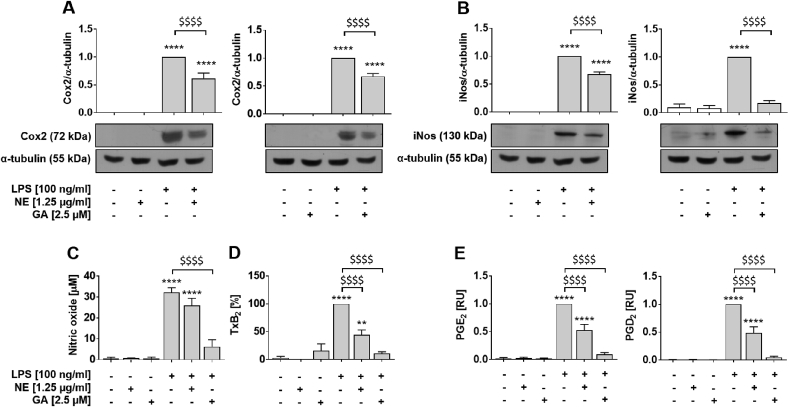
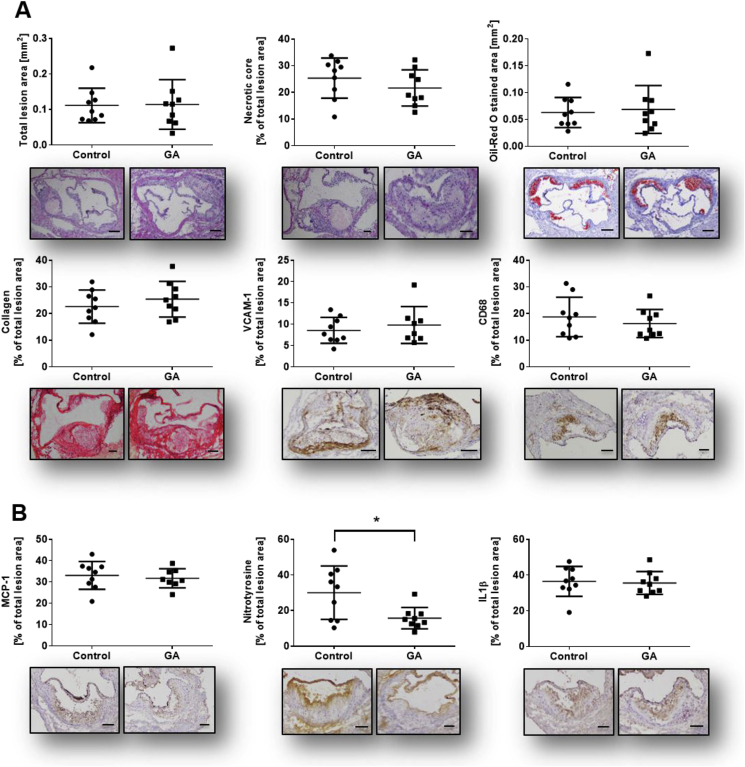
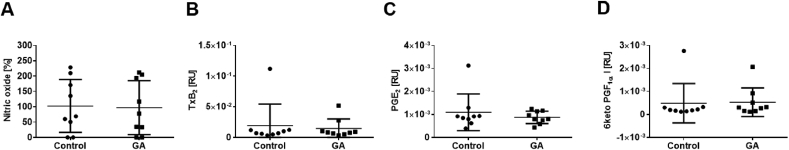
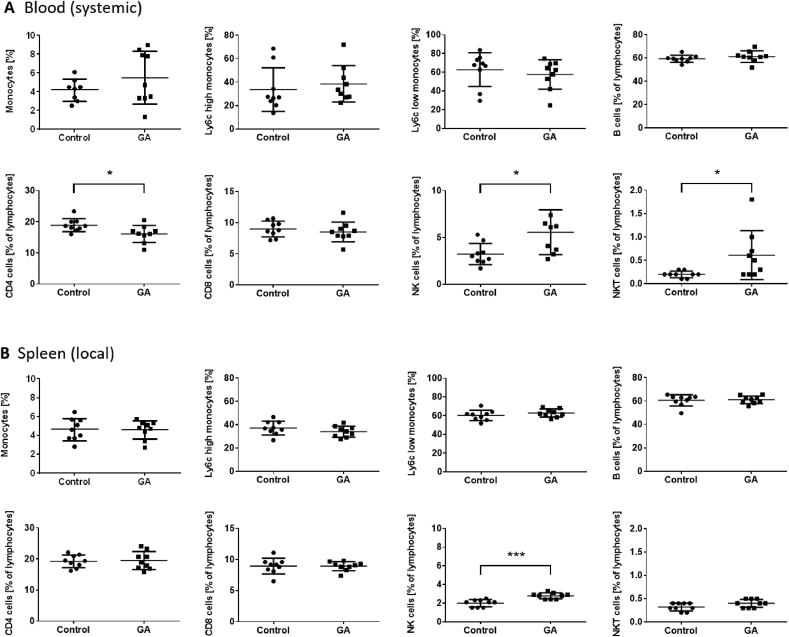
The plant Garcinia kola is used in African ethno-medicine to treat various oxidation- and inflammation-related diseases but its bioactive compounds are not well characterized. Garcinoic acid (GA) is one of the few phytochemicals that have been isolated from Garcinia kola. We investigated the anti-inflammatory potential of the methanol extract of Garcinia kola seeds (NE) and purified GA, as a major phytochemical in these seeds, in lipopolysaccharide (LPS)-activated mouse RAW264.7 macrophages and its anti-atherosclerotic potential in high fat diet fed ApoE-/- mice. This study outlines an optimized procedure for the extraction and purification of GA from Garcinia kola seeds with an increased yield and a purity of >99%. We found that LPS-induced upregulation of iNos and Cox2 expression, and the formation of the respective signaling molecules nitric oxide and prostanoids, were significantly diminished by both the NE and GA. In addition, GA treatment in mice decreased intra-plaque inflammation by attenuating nitrotyrosinylation. Further, modulation of lymphocyte sub-populations in blood and spleen have been detected, showing immune regulative properties of GA. Our study provides molecular insights into the anti-inflammatory activities of Garcinia kola and reveals GA as promising natural lead for the development of multi-target drugs to treat inflammation-driven diseases.
Keywords: Atherosclerosis; Garcinia kola seeds; Garcinoic acid; Inflammatory response; Macrophage activation.
Copyright © 2019 The Authors. Published by Elsevier B.V. All rights reserved.
Figures
References
-
- Iwu M.M., Duncan A.R., Okunji C.O. New antimicrobials of plant origin. Am. Soc. Hortic. Sci. Press. 1999:457–462.
-
- Hussain R.A., Owegby A.G., Parimoo P., Waterman P. G. Kolanone. A novel polyisoprenylated benzophenone with antimicrobial properties from the fruit of Garcinia kola. Planta Med. 1982;44:78–81. - PubMed
-
- Okoko T. In vitro antioxidant and free radical scavenging activities of Garcinia kola seeds. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2009;47:2620–2623. - PubMed
-
- Terashima K., Takaya Y., Niwa M. Powerful antioxidative agents based on garcinoic acid from Garcinia kola. Bioorg. Med. Chem. 2002;10:1619–1625. - PubMed
-
- Tchimene M.K. Bio-flavonoids and garcinoic acid from Garcinia kola seeds with promising local anesthetic potentials. Int. J. Pharmacogn. Phytochem. Res. 2015;7:764–767.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
