

VvmiR160s/VvARFs interaction and their spatio-temporal expression/cleavage products during GA-induced grape parthenocarpy
- PMID: 30898085
- PMCID: PMC6429806
- DOI: 10.1186/s12870-019-1719-9
VvmiR160s/VvARFs interaction and their spatio-temporal expression/cleavage products during GA-induced grape parthenocarpy
Abstract
Background: Grape (Vitis vinifera) is highly sensitive to gibberellin (GA), which effectively induce grape parthenocarpy. Studies showed that miR160s and their target AUXIN RESPONSIVE FACTOR (ARF) responding hormones are indispensable for various aspects of plant growth and development, but their functions in GA-induced grape parthenocarpy remain elusive.
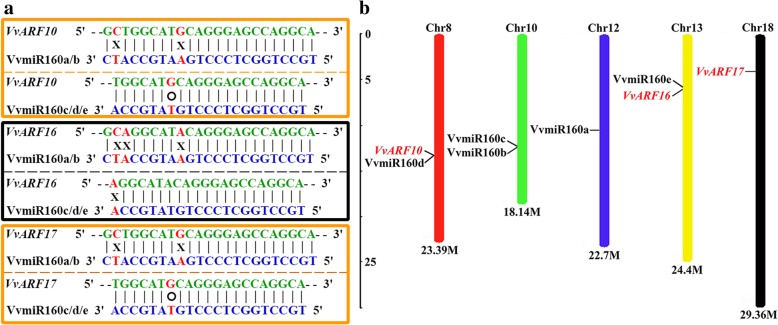
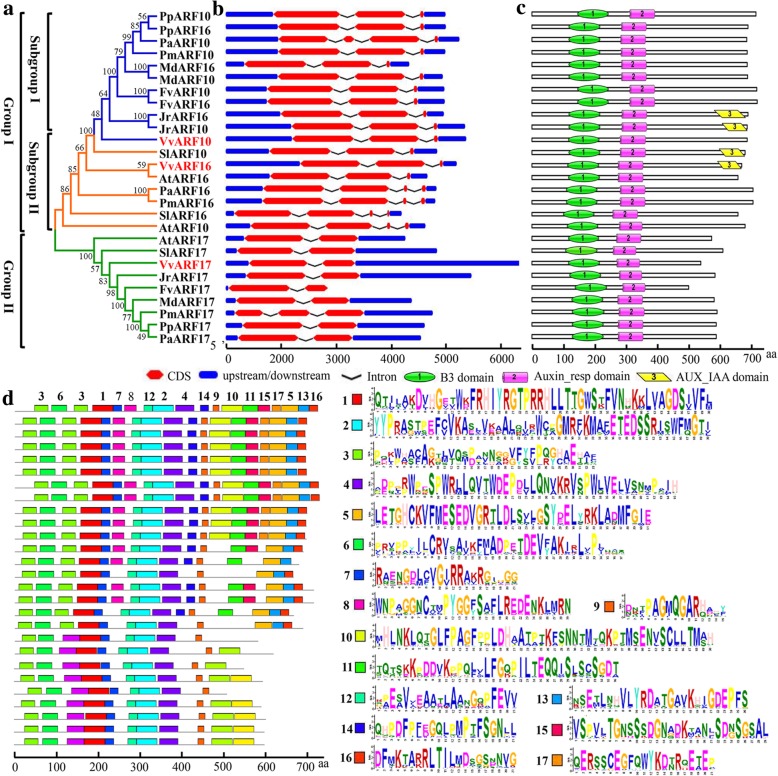
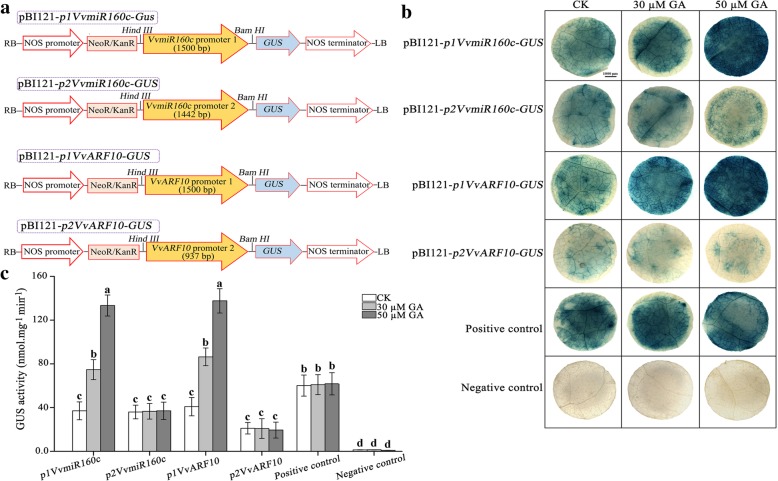
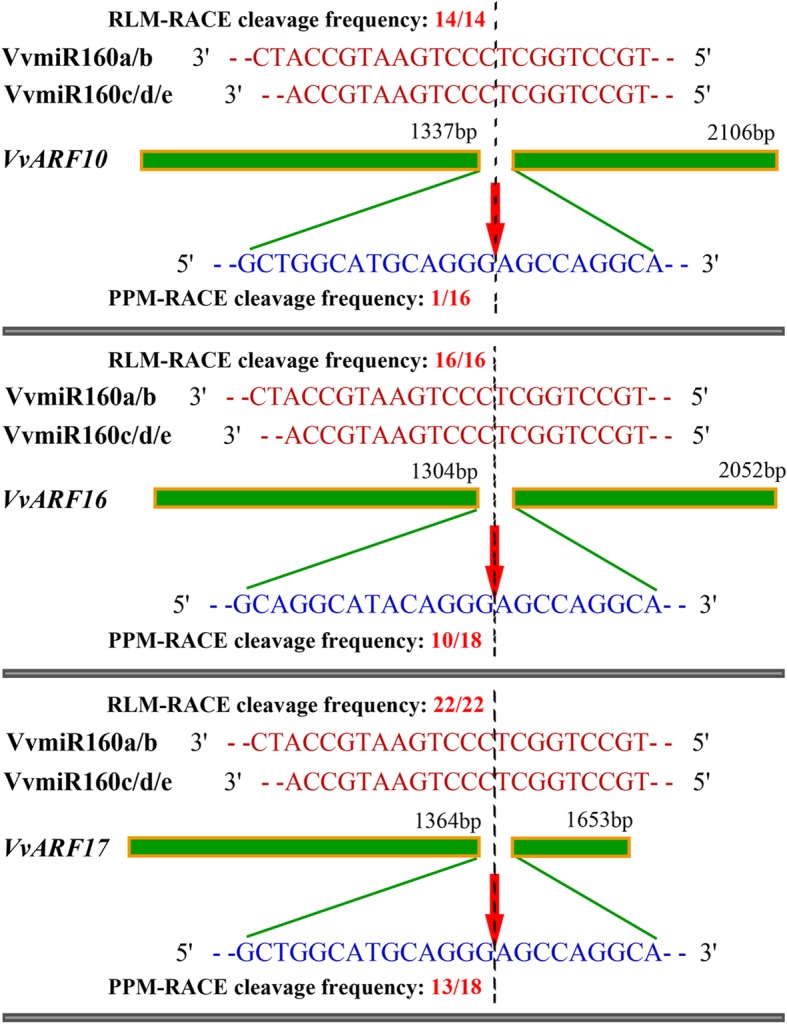
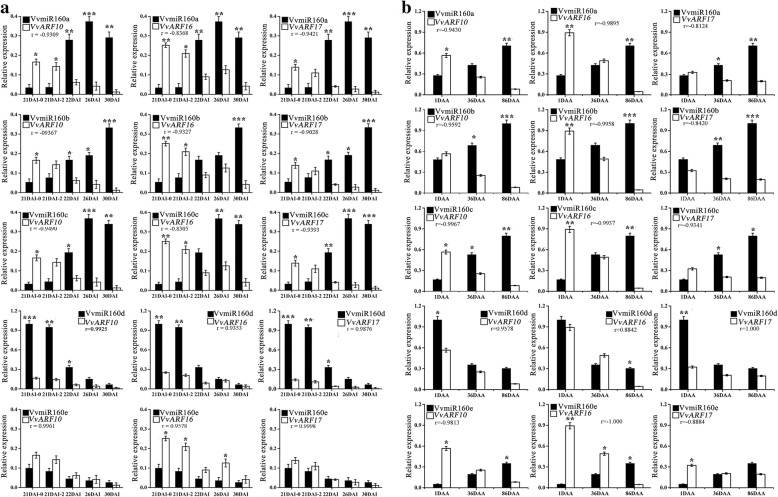
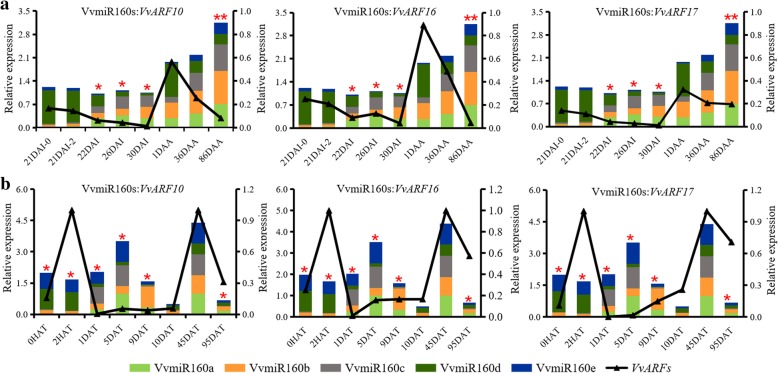
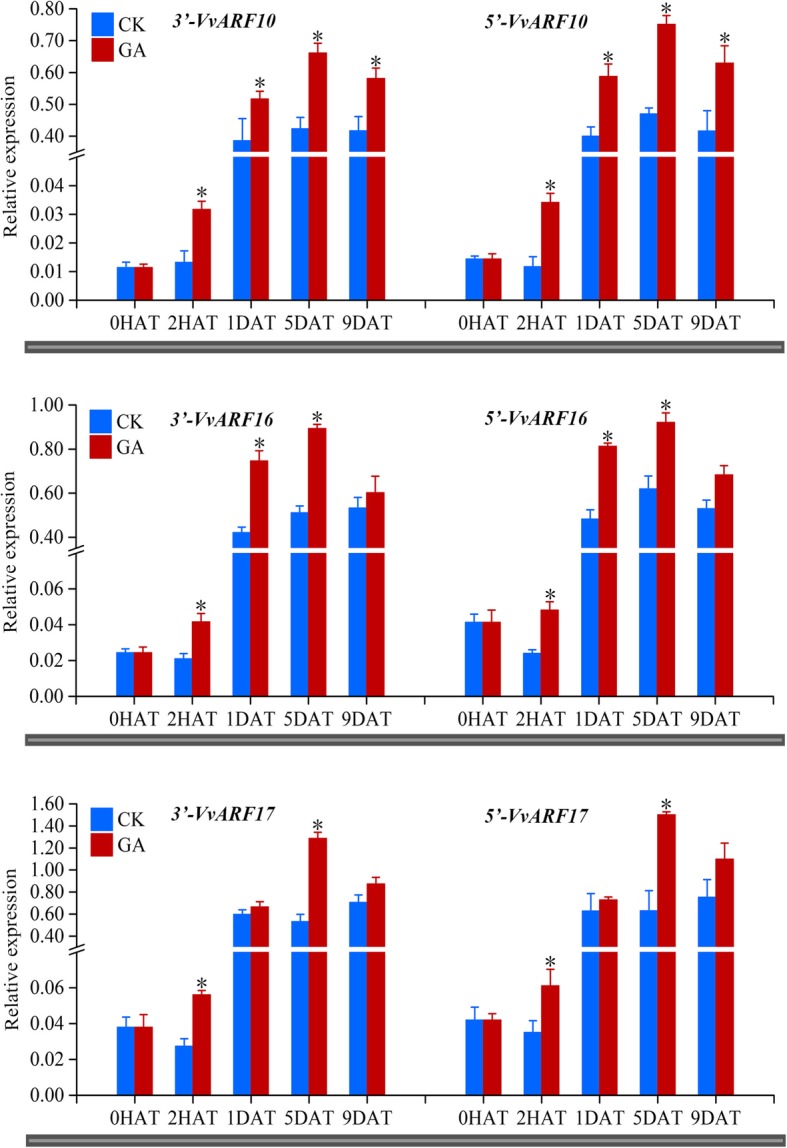
Results: In this study, the morphological changes during flower development in response to GA treatments were examined in the 'Rosario Bianco' cultivar. The precise sequences of VvmiR160a/b/c/d/e and their VvARF10/16/17 target genes were cloned, sequenced and characterized. The phylogenetic relationship and intron-exon structure of VvARFs and other ARF family members derived from different species were investigated. All VvmiR160s (except VvmiR160b) and VvARF10/16/17 had the common cis-elements responsive to GA, which support their function in GA-mediated grape parthenocarpy. The cleavage role of VvmiR160s-mediated VvARF10/16/17 was verified in grape flowers. Moreover, spatio-temporal expression analysis demonstrated that among VvmiR160 family, VvmiR160a/b/c highly expressed at late stage of flower/berry development, while VvARF10/16/17showed a reverse expression trend. Interestingly, GA exhibited a long-term effect through inducing the expression of VvmiR160a/b/c/e to increase their cleavage product accumulations from 5 to 9 days after treatment, but GA enhanced the expressions of VvARF10/16/17 only at short term. Pearson correlation analysis based on expression data revealed a negative correlation between VvmiR160a/b/c and VvARF10/16/17 in flowers not berries during GA-induced grape parthenocarpy.
Conclusions: This work demonstrated that the negative regulation of VvARF10/16/17 expression by VvmiR160a/b/c as key regulatory factors is critical for GA-mediated grape parthenocarpy, and provide significant implications for molecular breeding of high-quality seedless berry.
Keywords: Flower; Gibberellin; Grape; Parthenocarpy; VvARFs; VvmiR160s.
Conflict of interest statement
Ethics approval and consent to participate
The plants used in our study are not endangered species. Plant sample collection and transgenic plants were performed in accordance with the local legislation in China.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures




Similar articles
-
Spatio-temporal expression of miRNA159 family members and their GAMYB target gene during the modulation of gibberellin-induced grapevine parthenocarpy.J Exp Bot. 2018 Jun 27;69(15):3639-3650. doi: 10.1093/jxb/ery172. J Exp Bot. 2018. PMID: 29905866
-
Identification and Characterization of AUXIN Response Factor Gene Family Reveals Their Regulatory Network to Respond the Multi-Hormones Crosstalk during GA-Induced Grape Parthenocarpic Berry.Int J Mol Sci. 2022 Sep 21;23(19):11108. doi: 10.3390/ijms231911108. Int J Mol Sci. 2022. PMID: 36232409 Free PMC article.
-
Gibberellin Mediates VvmiR397a-VvLAC4 via VvSLR1-VvWRKY26 Cascade Signal to Repress the Seed-Stone Development During GA-Induced Grape Parthenocarpy.Plant Cell Environ. 2025 Aug;48(8):5930-5949. doi: 10.1111/pce.15570. Epub 2025 Apr 23. Plant Cell Environ. 2025. PMID: 40269609
-
Trans-acting small interfering RNA4: key to nutraceutical synthesis in grape development?Trends Plant Sci. 2013 Nov;18(11):601-10. doi: 10.1016/j.tplants.2013.07.006. Epub 2013 Aug 28. Trends Plant Sci. 2013. PMID: 23993483 Free PMC article. Review.
-
The making of virgin fruit: the molecular and genetic basis of parthenocarpy.J Exp Bot. 2018 Feb 23;69(5):955-962. doi: 10.1093/jxb/erx446. J Exp Bot. 2018. PMID: 29325151 Free PMC article. Review.
Cited by
-
Roles of the SPL gene family and miR156 in the salt stress responses of tamarisk (Tamarix chinensis).BMC Plant Biol. 2019 Aug 22;19(1):370. doi: 10.1186/s12870-019-1977-6. BMC Plant Biol. 2019. PMID: 31438851 Free PMC article.
-
Convergent domestication of bitter apples and pears by selecting mutations of MYB transcription factors to reduce proanthocyanidin levels.Mol Hortic. 2025 Sep 4;5(1):51. doi: 10.1186/s43897-025-00173-z. Mol Hortic. 2025. PMID: 40903764 Free PMC article.
-
Characterization of VvmiR166s-Target Modules and Their Interaction Pathways in Modulation of Gibberellic-Acid-Induced Grape Seedless Berries.Int J Mol Sci. 2023 Nov 14;24(22):16279. doi: 10.3390/ijms242216279. Int J Mol Sci. 2023. PMID: 38003470 Free PMC article.
-
Identification of miRNAs-mediated seed and stone-hardening regulatory networks and their signal pathway of GA-induced seedless berries in grapevine (V. vinifera L.).BMC Plant Biol. 2021 Sep 29;21(1):442. doi: 10.1186/s12870-021-03188-y. BMC Plant Biol. 2021. PMID: 34587914 Free PMC article.
-
Effects of plant growth regulator (2,4-D) treatments on fruit development and chemical content in the kiwifruit plant (Actinidia chinensis).BMC Plant Biol. 2025 Aug 14;25(1):1073. doi: 10.1186/s12870-025-07138-w. BMC Plant Biol. 2025. PMID: 40813636 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
