

A complex of novel protease inhibitor, ovostatin homolog, with its cognate proteases in immature mice uterine luminal fluid
- PMID: 30899053
- PMCID: PMC6428836
- DOI: 10.1038/s41598-019-41426-4
A complex of novel protease inhibitor, ovostatin homolog, with its cognate proteases in immature mice uterine luminal fluid
Abstract
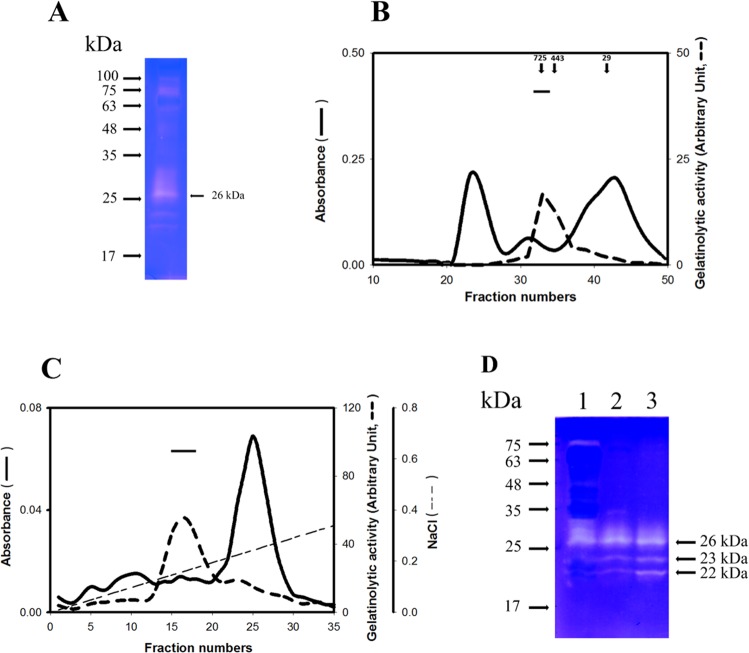
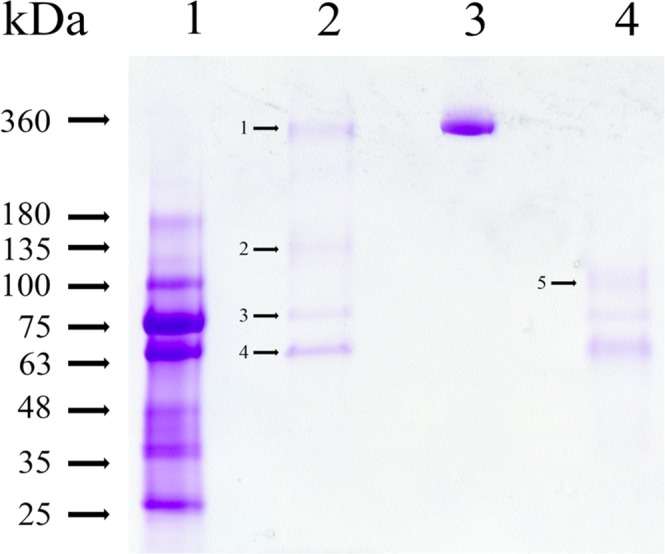
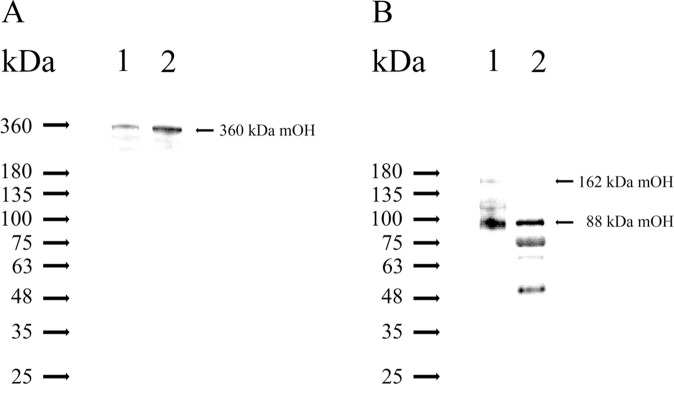
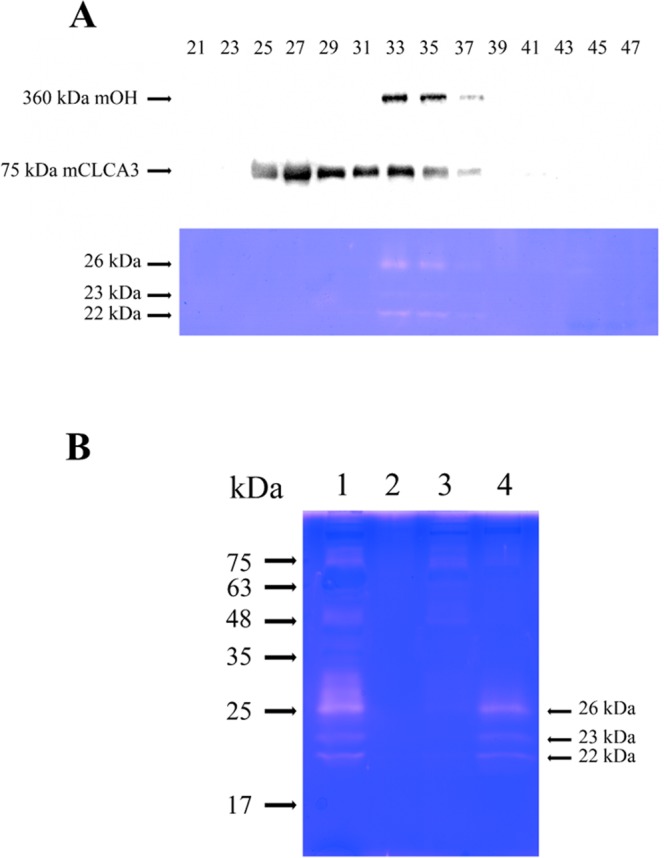
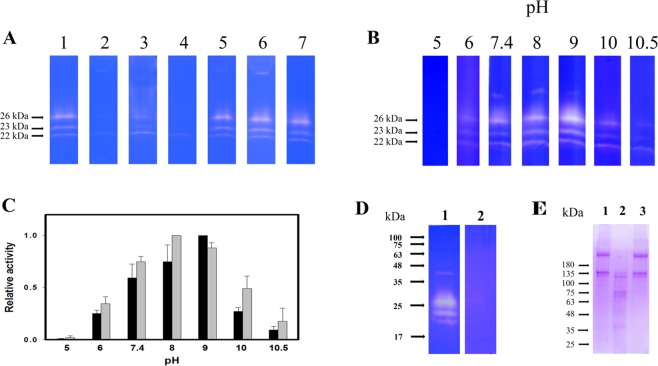
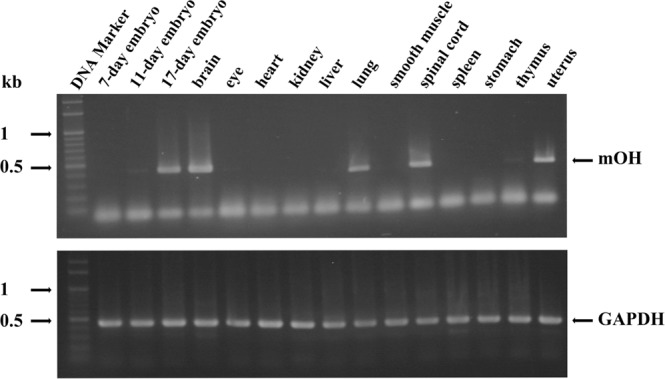
A predominant gelatinolytic enzyme with approximately 26 kDa was observed in gelatin zymogram of immature mice uterine luminal fluid (ULF). Size exclusion analysis revealed that the native size of this enzyme was close to that of human α2-macroglobulin (α2-MG), a 725 kDa protein. This large protease was isolated by a series of chromatographic steps on the Sephacryl S-400 and DEAE-Sepharose columns. The results from gelatin zymography and SDS-PAGE analysis supported that this large protease consists of gelatinolytic enzyme and a 360 kDa protein. Through tandem mass spectrometry analysis followed by MASCOT database search, the 360 kDa protein was identified as ovostatin homolog (accession: NP_001001179.2) assigned as a homolog of chicken ovostatin, a protease inhibitor. The co-fractionation analysis by gel filtration and mouse ovostatin homolog (mOH) co-immunoprecipitation experiments demonstrated that the mOH formed a complex with three gelatinolytic enzymes in immature mice ULF. Substrate zymography analysis revealed that the mOH-associated gelatinolytic enzymes were suitable to digest type I collagen rather than type IV collagen. In addition, the refolded mOH-associated 26 kDa gelatinolytic enzyme displayed the type I collagen-digesting activity in the assay, but the other two enzymes did not have this function. RT-PCR analysis showed that mOH gene was abundantly expressed in brain, spinal cord, lung, uterus, and in 17-day embryo. Taken together, our data suggest that mOH/cognate protease system may play a potential role in regulation of tissue remodeling and fetal development.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Evans GS, Gibson DF, Roberts SA, Hind TM, Potten CS. Proliferative changes in the genital tissue of female mice during the oestrous cycle. Cell and Tissue Kinetics. 1990;23:619–635. - PubMed
-
- Rugh, R. Reproductive Systems of Adult Mice, New York: Oxford University Press. (1990).
-
- Ricci S, D’Esposito V, Oriente F, Formisano P, Di Carlo A. Substrate-zymography: a still worthwhile method for gelatinases analysis in biological sample. Clin Chem Lab Med. 2016;54:1281–1290. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
