

The Small and Dynamic Pre-primed Pool at the Release Site; A Useful Concept to Understand Release Probability and Short-Term Synaptic Plasticity?
- PMID: 30899219
- PMCID: PMC6416800
- DOI: 10.3389/fnsyn.2019.00007
The Small and Dynamic Pre-primed Pool at the Release Site; A Useful Concept to Understand Release Probability and Short-Term Synaptic Plasticity?
Abstract
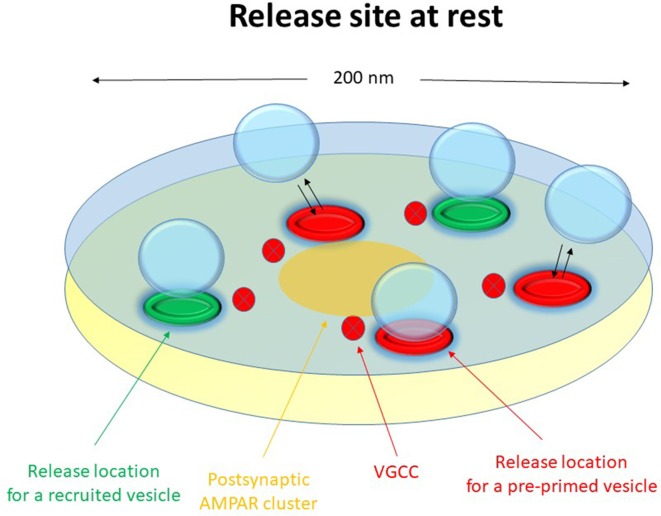
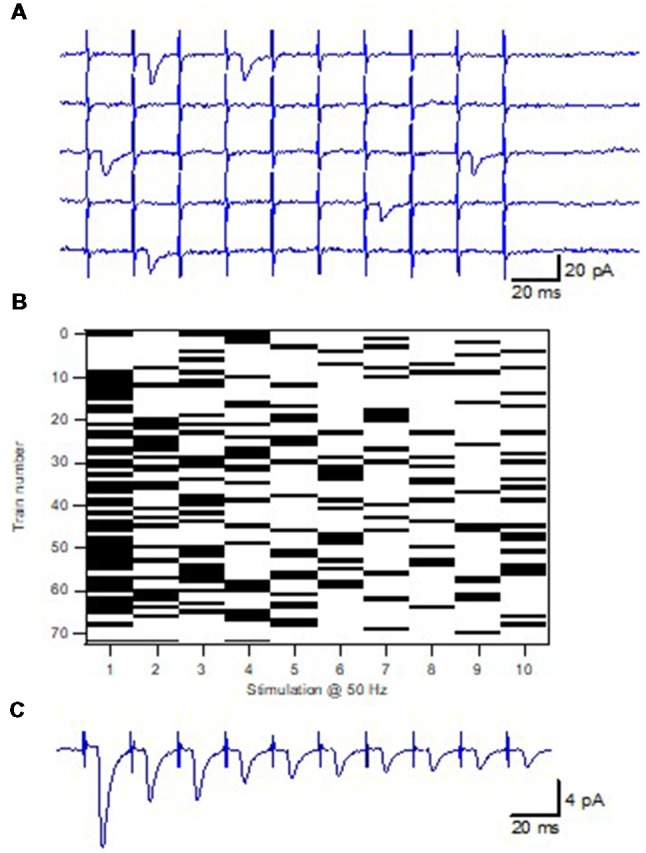
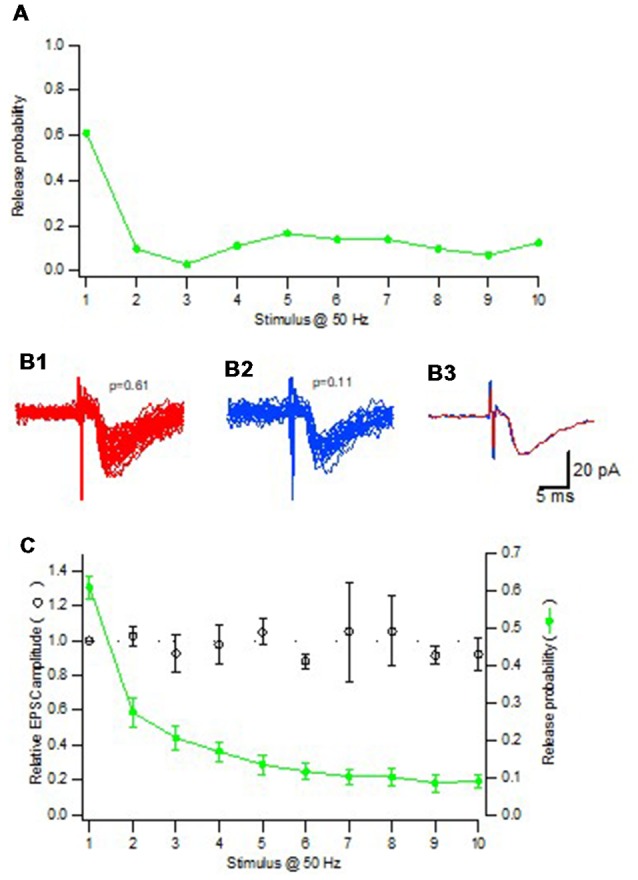
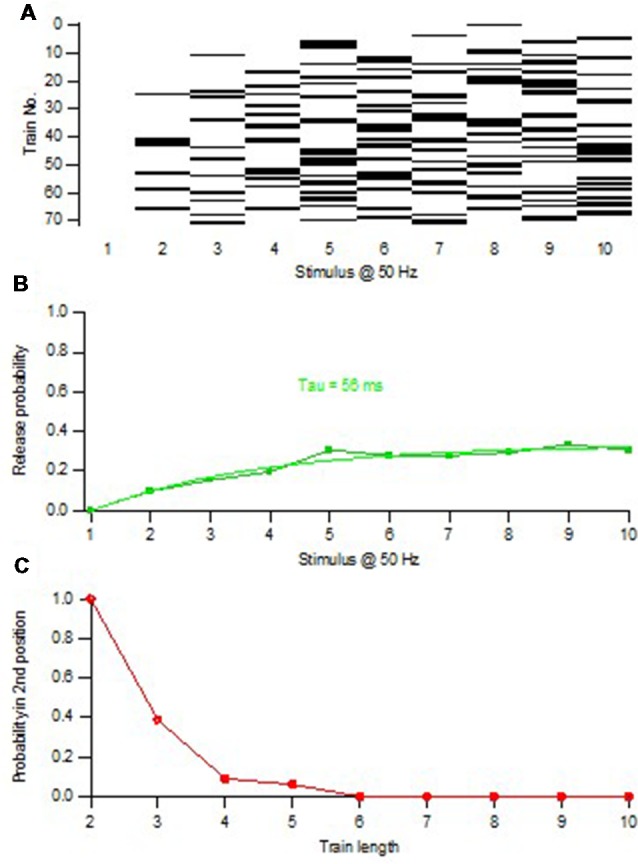
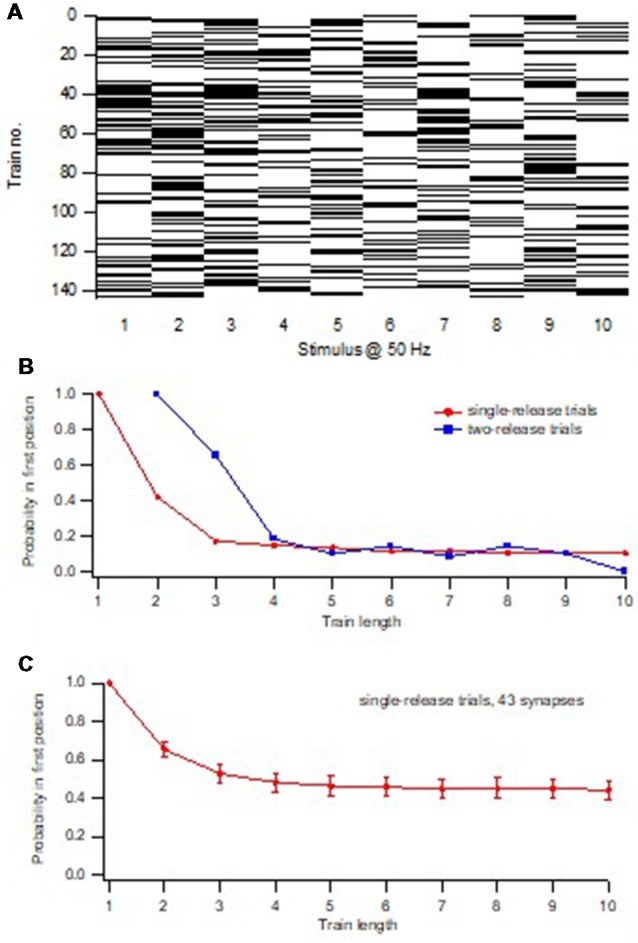
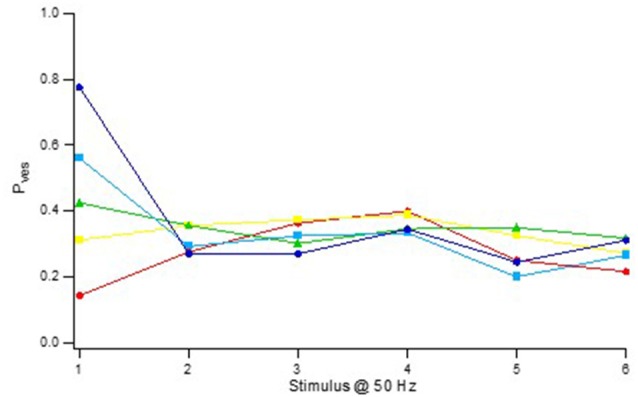
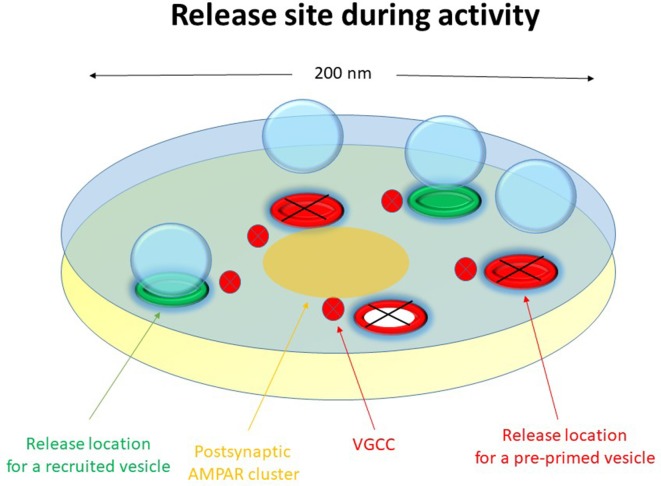
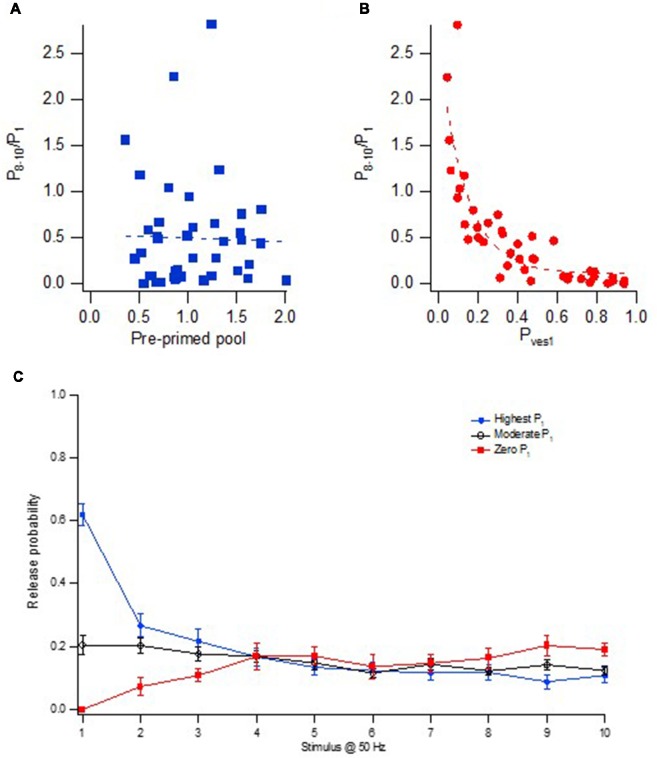
Advanced imaging techniques have revealed that synapses contain nanomodules in which pre- and post-synaptic molecules are brought together to form an integrated subsynaptic component for vesicle release and transmitter reception. Based on data from an electrophysiological study of ours in which release from synapses containing a single nanomodule was induced by brief 50 Hz trains using minimal stimulation, and on data from such imaging studies, we present a possible modus operandi of such a nanomodule. We will describe the techniques and tools used to obtain and analyze the electrophysiological data from single CA3-CA1 hippocampal synapses from the neonatal rat brain. This analysis leads to the proposal that a nanomodule, despite containing a number of release locations, operates as a single release site, releasing at most a single vesicle at a time. In this nanomodule there appears to be two separate sets of release locations, one set that is responsible for release in response to the first few action potentials and another set that produces the release thereafter. The data also suggest that vesicles at the first set of release locations are primed by synaptic inactivity lasting seconds, this synaptic inactivity also resulting in a large heterogeneity in the values for vesicle release probability among the synapses. The number of vesicles being primed at this set of release locations prior to the arrival of an action potential is small (0-3) and varies from train to train. Following the first action potential, this heterogeneity in vesicle release probability largely vanishes in a release-independent manner, shaping a variation in paired-pulse plasticity among the synapses. After the first few action potentials release is produced from the second set of release locations, and is given by vesicles that have been recruited after the onset of synaptic activity. This release depends on the number of such release locations and the recruitment to such a location. The initial heterogeneity in vesicle release probability, its disappearance after a single action potential, and variation in the recruitment to the second set of release locations are instrumental in producing the heterogeneity in short-term synaptic plasticity among these synapses, and can be seen as means to create differential dynamics within a synapse population.
Keywords: glutamate; hippocampus; nanomodule; plasticity; release probability; release site; synapse; vesicle.
Figures
Similar articles
-
Vesicle release probability and pre-primed pool at glutamatergic synapses in area CA1 of the rat neonatal hippocampus.J Physiol. 2001 Mar 1;531(Pt 2):481-93. doi: 10.1111/j.1469-7793.2001.0481i.x. J Physiol. 2001. PMID: 11230520 Free PMC article.
-
Factors explaining heterogeneity in short-term synaptic dynamics of hippocampal glutamatergic synapses in the neonatal rat.J Physiol. 2001 Nov 15;537(Pt 1):141-9. doi: 10.1111/j.1469-7793.2001.0141k.x. J Physiol. 2001. PMID: 11711568 Free PMC article.
-
Mechanisms of target-cell specific short-term plasticity at Schaffer collateral synapses onto interneurones versus pyramidal cells in juvenile rats.J Physiol. 2005 Nov 1;568(Pt 3):815-40. doi: 10.1113/jphysiol.2005.093948. Epub 2005 Aug 18. J Physiol. 2005. PMID: 16109728 Free PMC article.
-
Multiple vesicle recycling pathways in central synapses and their impact on neurotransmission.J Physiol. 2007 Dec 15;585(Pt 3):669-79. doi: 10.1113/jphysiol.2007.137745. Epub 2007 Aug 9. J Physiol. 2007. PMID: 17690145 Free PMC article. Review.
-
[Short term synaptic plasticity].Rev Neurol. 2002 Mar 16-31;34(6):593-9. Rev Neurol. 2002. PMID: 12040507 Review. Spanish.
Cited by
-
Zebrafish neuromuscular junction: The power of N.Neurosci Lett. 2019 Nov 20;713:134503. doi: 10.1016/j.neulet.2019.134503. Epub 2019 Sep 23. Neurosci Lett. 2019. PMID: 31557523 Free PMC article. Review.
-
A sequential two-step priming scheme reproduces diversity in synaptic strength and short-term plasticity.Proc Natl Acad Sci U S A. 2022 Aug 23;119(34):e2207987119. doi: 10.1073/pnas.2207987119. Epub 2022 Aug 15. Proc Natl Acad Sci U S A. 2022. PMID: 35969787 Free PMC article.
-
Rapid regulation of vesicle priming explains synaptic facilitation despite heterogeneous vesicle:Ca2+ channel distances.Elife. 2020 Feb 20;9:e51032. doi: 10.7554/eLife.51032. Elife. 2020. PMID: 32077852 Free PMC article.
-
Interpretation of presynaptic phenotypes of synaptic plasticity in terms of a two-step priming process.J Gen Physiol. 2024 Jan 1;156(1):e202313454. doi: 10.1085/jgp.202313454. Epub 2023 Dec 19. J Gen Physiol. 2024. PMID: 38112713 Free PMC article.
-
Elevated synaptic vesicle release probability in synaptophysin/gyrin family quadruple knockouts.Elife. 2019 May 15;8:e40744. doi: 10.7554/eLife.40744. Elife. 2019. PMID: 31090538 Free PMC article.
References
Publication types
LinkOut - more resources
Full Text Sources
Miscellaneous