

Astrogliogenesis in human fetal brain: complex spatiotemporal immunoreactivity patterns of GFAP, S100, AQP4 and YKL-40
- PMID: 30901080
- PMCID: PMC6704246
- DOI: 10.1111/joa.12948
Astrogliogenesis in human fetal brain: complex spatiotemporal immunoreactivity patterns of GFAP, S100, AQP4 and YKL-40
Abstract
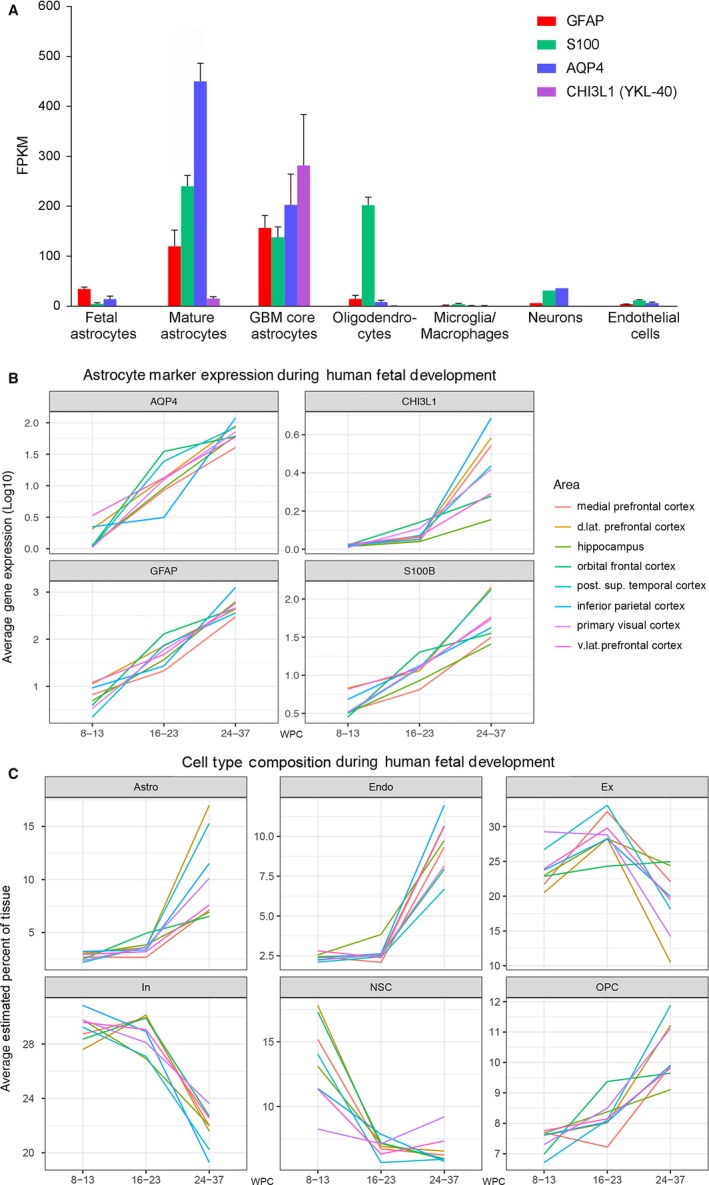
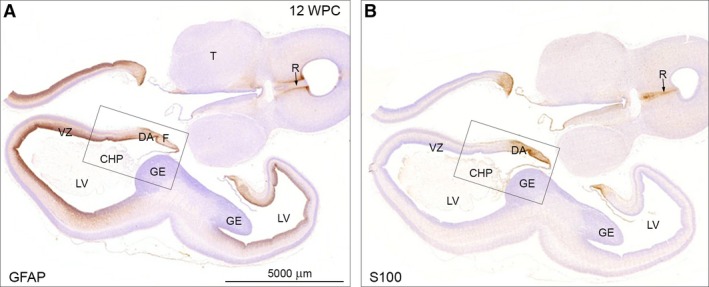
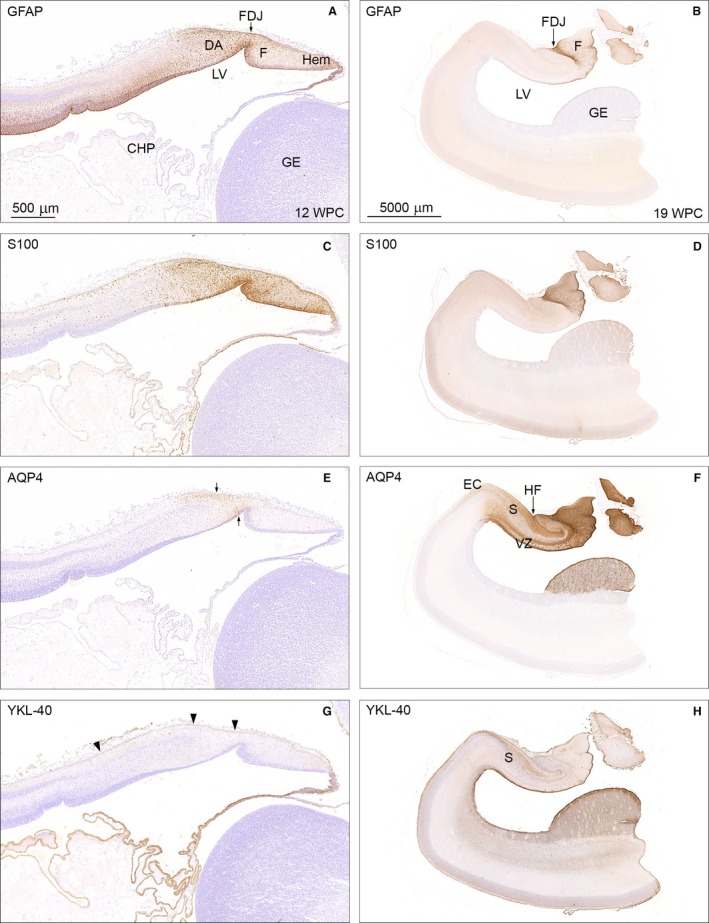
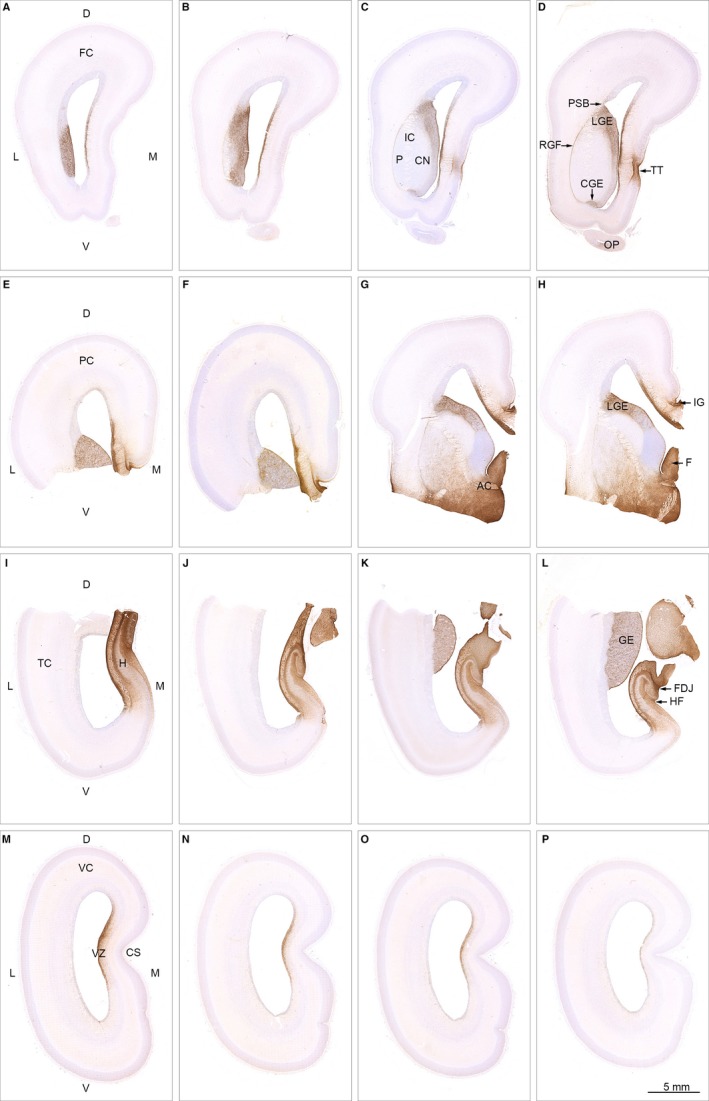
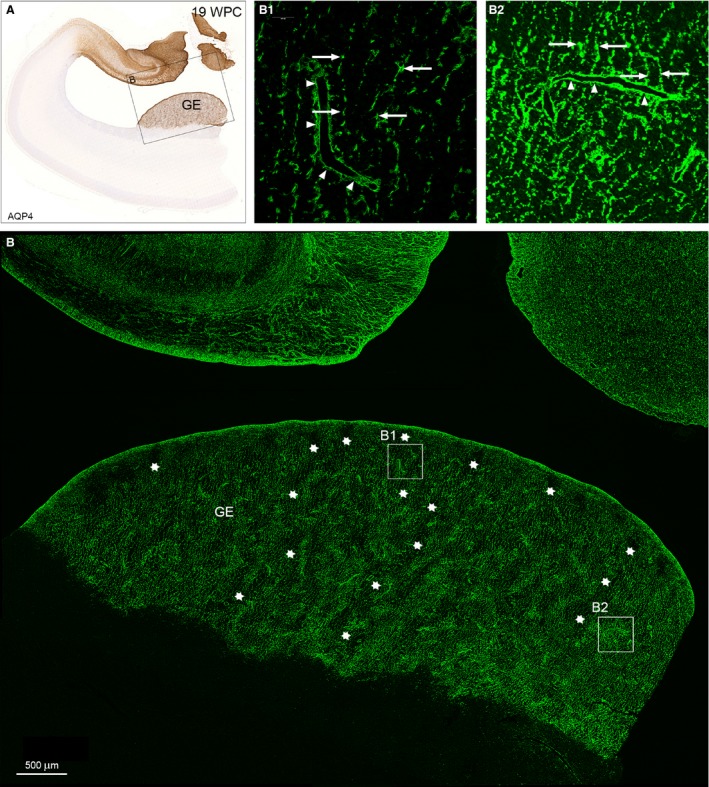
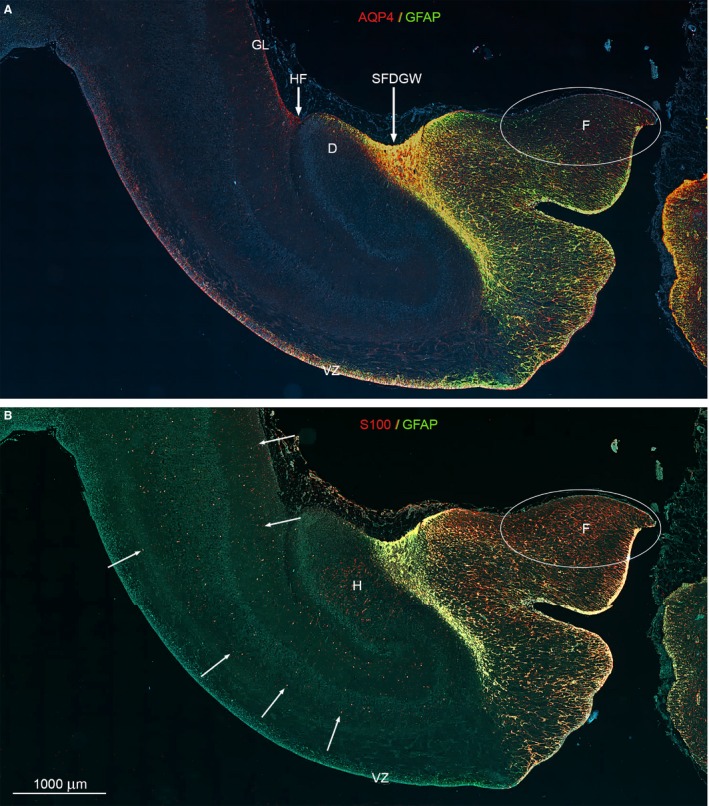
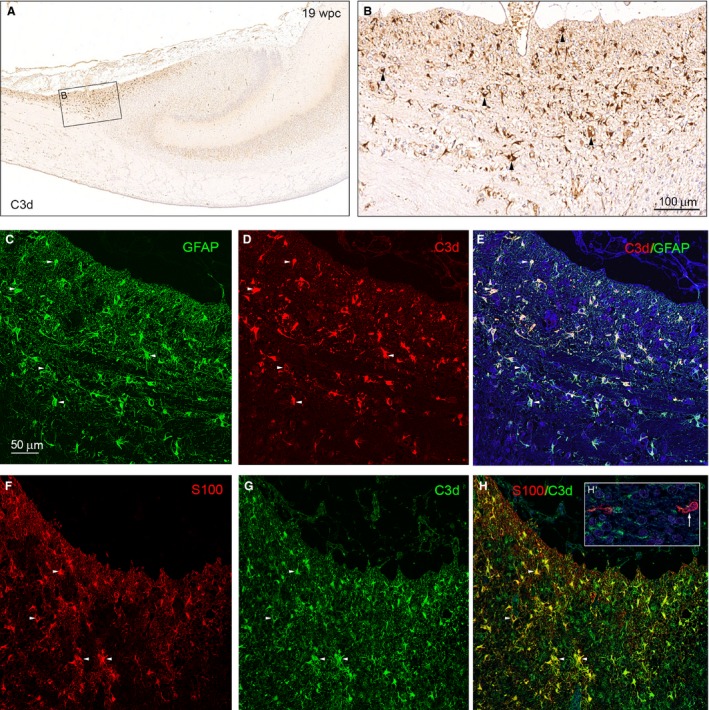
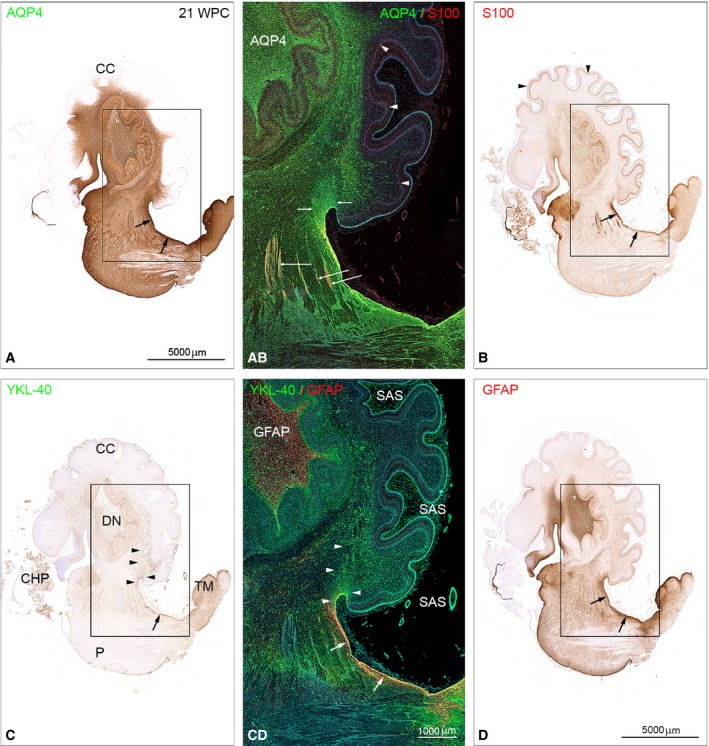
The astroglial lineage consists of heterogeneous cells instrumental for normal brain development, function and repair. Unfortunately, this heterogeneity complicates research in the field, which suffers from lack of truly specific and sensitive astroglial markers. Nevertheless, single astroglial markers are often used to describe astrocytes in different settings. We therefore investigated and compared spatiotemporal patterns of immunoreactivity in developing human brain from 12 to 21 weeks post conception and publicly available RNA expression data for four established and potential astroglial markers - GFAP, S100, AQP4 and YKL-40. In the hippocampal region, we also screened for C3, a complement component highly expressed in A1-reactive astrocytes. We found diverging partly overlapping patterns of the established astroglial markers GFAP, S100 and AQP4, confirming that none of these markers can fully describe and discriminate different developmental forms and subpopulations of astrocytes in human developing brain, although AQP4 seems to be the most sensitive and specific marker for the astroglial lineage at midgestation. AQP4 characterizes a brain-wide water transport system in cerebral cortex with regional differences in immunoreactivity at midgestation. AQP4 distinguishes a vast proportion of astrocytes and subpopulations of radial glial cells destined for the astroglial lineage, including astrocytes determined for the future glia limitans and apical truncated radial glial cells in ganglionic eminences, devoid of GFAP and S100. YKL-40 and C3d, previously found in reactive astrocytes, stain different subpopulations of astrocytes/astroglial progenitors in developing hippocampus at midgestation and may characterize specific subpopulations of 'developmental astrocytes'. Our results clearly reflect that lack of pan-astrocytic markers necessitates the consideration of time, region, context and aim when choosing appropriate astroglial markers.
Keywords: astrocytes; astrogliogenesis; glial markers; human brain development; radial glial cells.
© 2019 Anatomical Society.
Conflict of interest statement
The authors have no conflict of interest to declare.
Figures
References
-
- Abbott NJ, Ronnback L, Hansson E (2006) Astrocyte‐endothelial interactions at the blood‐brain barrier. Nat Rev Neurosci 7, 41–53. - PubMed
-
- Abbott NJ, Pizzo ME, Preston JE, et al. (2018) The role of brain barriers in fluid movement in the CNS: is there a ‘glymphatic’ system? Acta Neuropathol 135, 387–407. - PubMed
-
- Abraham H, Perez‐Garcia CG, Meyer G (2004) p73 and Reelin in Cajal‐Retzius cells of the developing human hippocampal formation. Cereb Cortex 14, 484–495. - PubMed
-
- Abraham H, Vincze A, Jewgenow I, et al. (2010) Myelination in the human hippocampal formation from midgestation to adulthood. Int J Dev Neurosci 28, 401–410. - PubMed
-
- Allen NJ (2013) Role of glia in developmental synapse formation. Curr Opin Neurobiol 23, 1027–1033. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
