

A blueprint of mammalian cortical connectomes
- PMID: 30901324
- PMCID: PMC6456226
- DOI: 10.1371/journal.pbio.2005346
A blueprint of mammalian cortical connectomes
Abstract
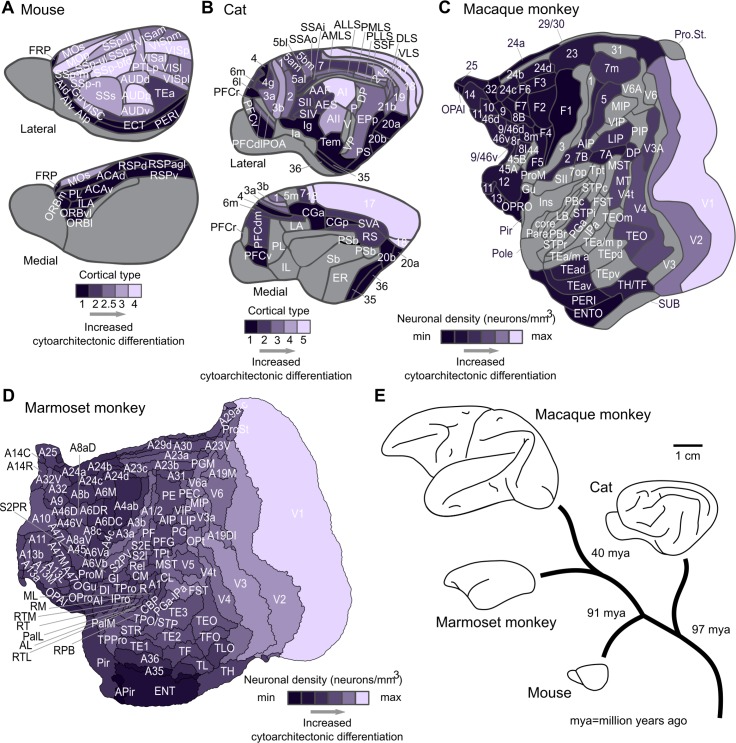
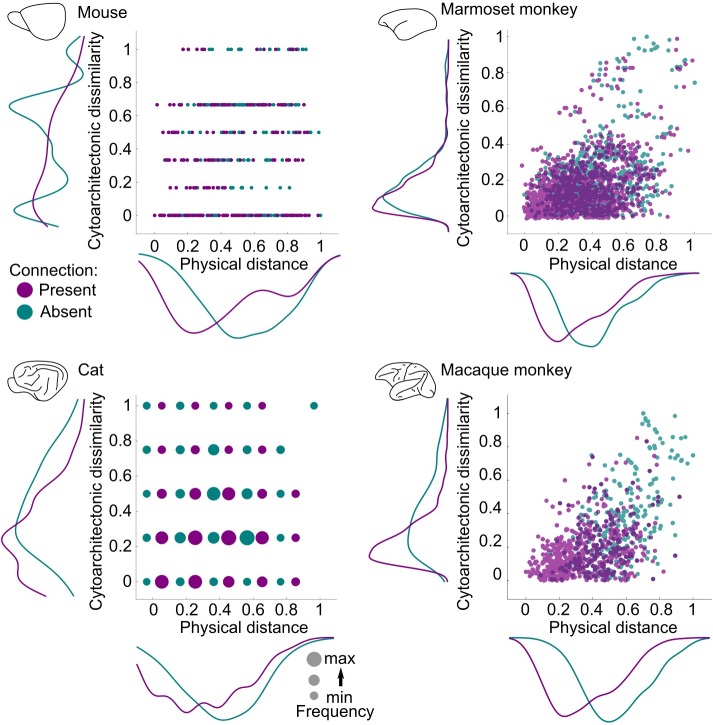
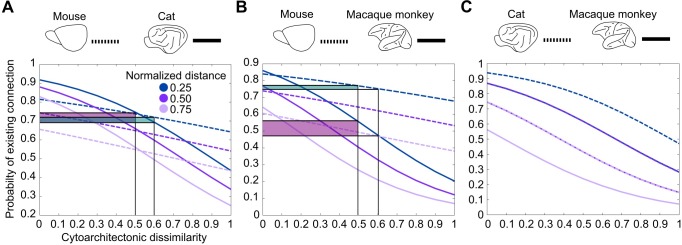
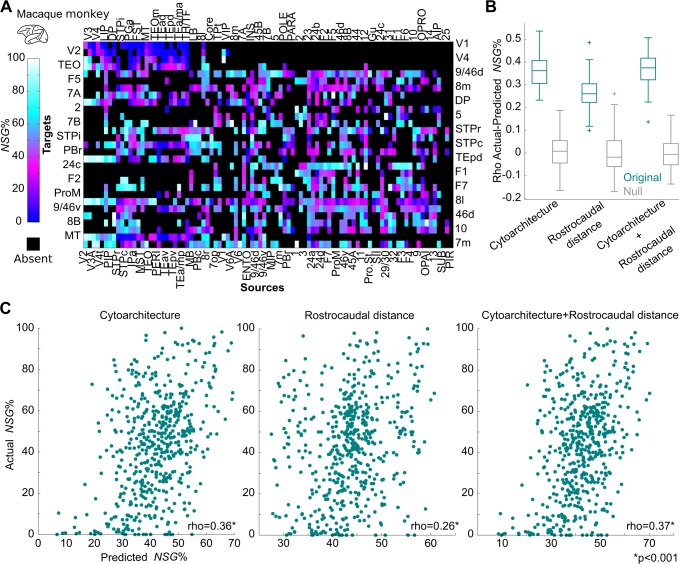
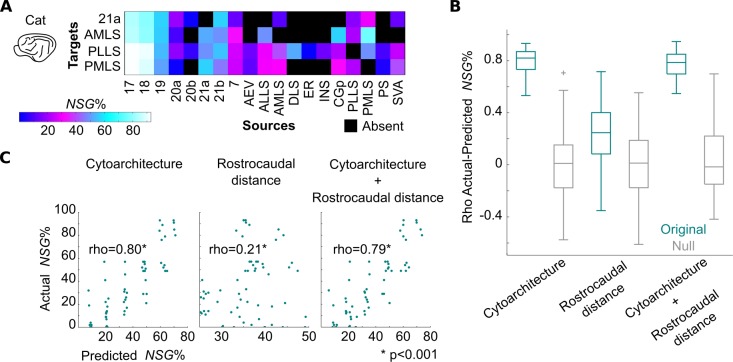
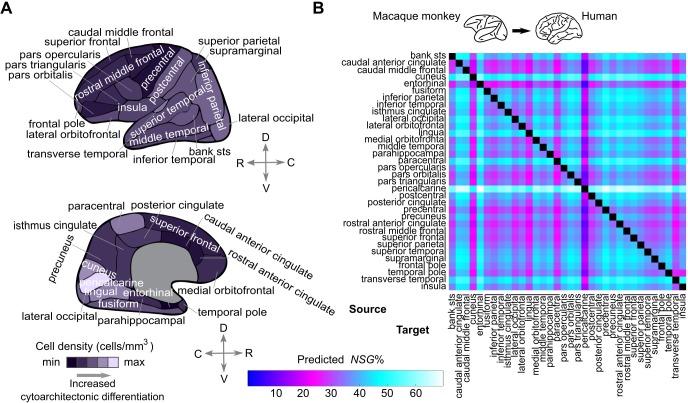
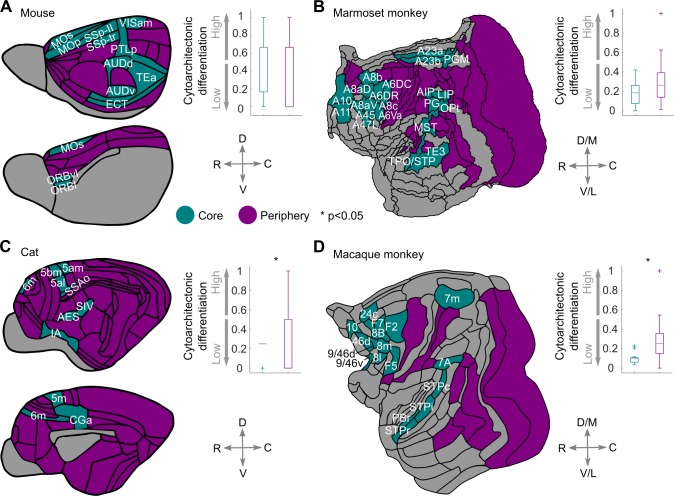
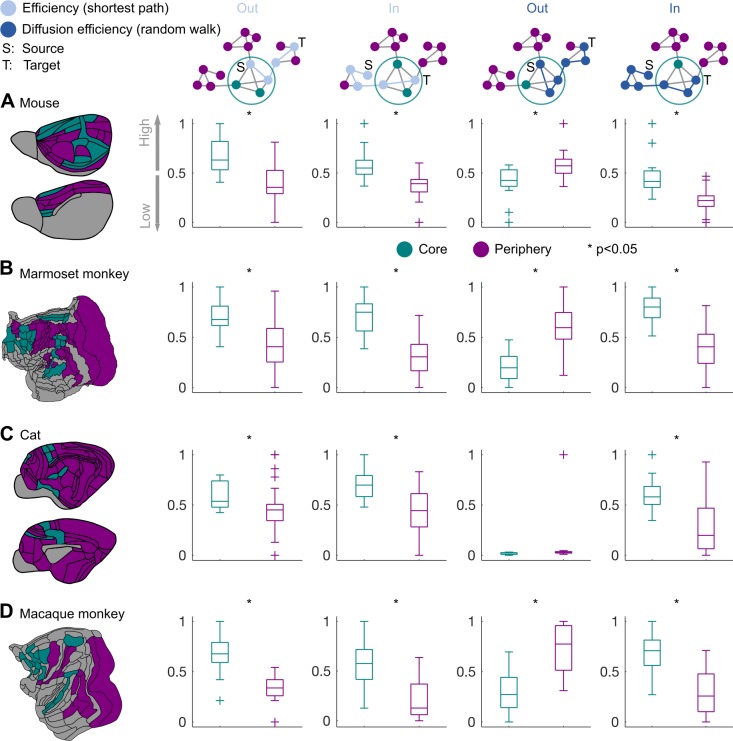
The cerebral cortex of mammals exhibits intricate interareal wiring. Moreover, mammalian cortices differ vastly in size, cytological composition, and phylogenetic distance. Given such complexity and pronounced species differences, it is a considerable challenge to decipher organizational principles of mammalian connectomes. Here, we demonstrate species-specific and species-general unifying principles linking the physical, cytological, and connectional dimensions of architecture in the mouse, cat, marmoset, and macaque monkey. The existence of connections is related to the cytology of cortical areas, in addition to the role of physical distance, but this relation is attenuated in mice and marmoset monkeys. The cytoarchitectonic cortical gradients, and not the rostrocaudal axis of the cortex, are closely linked to the laminar origin of connections, a principle that allows the extrapolation of this connectional feature to humans. Lastly, a network core, with a central role under different modes of network communication, characterizes all cortical connectomes. We observe a displacement of the network core in mammals, with a shift of the core of cats and macaque monkeys toward the less neuronally dense areas of the cerebral cortex. This displacement has functional ramifications but also entails a potential increased degree of vulnerability to pathology. In sum, our results sketch out a blueprint of mammalian connectomes consisting of species-specific and species-general links between the connectional, physical, and cytological dimensions of the cerebral cortex, possibly reflecting variations and persistence of evolutionarily conserved mechanisms and cellular phenomena. Our framework elucidates organizational principles that encompass but also extend beyond the wiring economy principle imposed by the physical embedding of the cerebral cortex.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Comment in
-
Evolution, development, and organization of the cortical connectome.PLoS Biol. 2019 May 10;17(5):e3000259. doi: 10.1371/journal.pbio.3000259. eCollection 2019 May. PLoS Biol. 2019. PMID: 31075099 Free PMC article.
References
-
- Braitenberg V, Schüz A. Anatomy of the Cortex-Statistics and Geometry. 1st ed London, New York: Springer; 1991.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous
