

Pirt deficiency has subtle female-specific effects on energy and glucose metabolism in mice
- PMID: 30902502
- PMCID: PMC6479763
- DOI: 10.1016/j.molmet.2019.02.011
Pirt deficiency has subtle female-specific effects on energy and glucose metabolism in mice
Erratum in
-
Corrigendum to "Pirt deficiency has subtle female-specific effects on energy and glucose metabolism in mice" [Molecular Metabolism 23 (2019) 75-81].Mol Metab. 2019 Dec;30:264. doi: 10.1016/j.molmet.2019.11.002. Mol Metab. 2019. PMID: 31767177 Free PMC article. No abstract available.
Abstract
Objective: The contribution of brown adipose tissue (BAT) to adult human metabolic control is a topic of ongoing investigation. In context, understanding the cellular events leading to BAT uncoupling, heat production, and energy expenditure is anticipated to produce significant insight into this endeavor. The phosphoinositide interacting regulator of transient receptor potentials (Pirt) was recently put forward as a key protein regulating cold sensing downstream of the transient receptor potential melastatin 8 (TRPM8). Notably, TRPM8 has been identified as a non-canonical regulator of BAT thermogenesis. The aim of this investigation was to delineate the role of Pirt in energy homeostasis and glucose metabolism - and the possible involvement of Pirt in TRPM8-elicited energy expenditure.
Methods: To this end, we metabolically phenotyped male and female Pirt deficient (Pirt-/-) mice exposed to a low-fat chow diet or to a high-fat, high-sugar (HFHS) diet.
Results: We identified that chow-fed female Pirt-/- mice have an increased susceptibility to develop obesity and glucose intolerance. This effect is abrogated when the mice are exposed to a HFHS diet. Conversely, Pirt-/- male mice display no metabolic phenotype on either diet relative to wild-type (WT) control mice. Finally, we observed that Pirt is dispensable for TRPM8-evoked energy expenditure.
Conclusion: We here report subtle metabolic abnormalities in female, but not male, Pirt-/- mice. Future studies are required to tease out if metabolic stressors beyond dietary interventions, e.g. temperature fluctuations, are interacting with Pirt-signaling and metabolic control in a sex-specific fashion.
Keywords: Body weight; Brown adipose tissue; Energy metabolism; Sex differences; Signaling molecule; TRPM8.
Copyright © 2019 The Authors. Published by Elsevier GmbH.. All rights reserved.
Figures
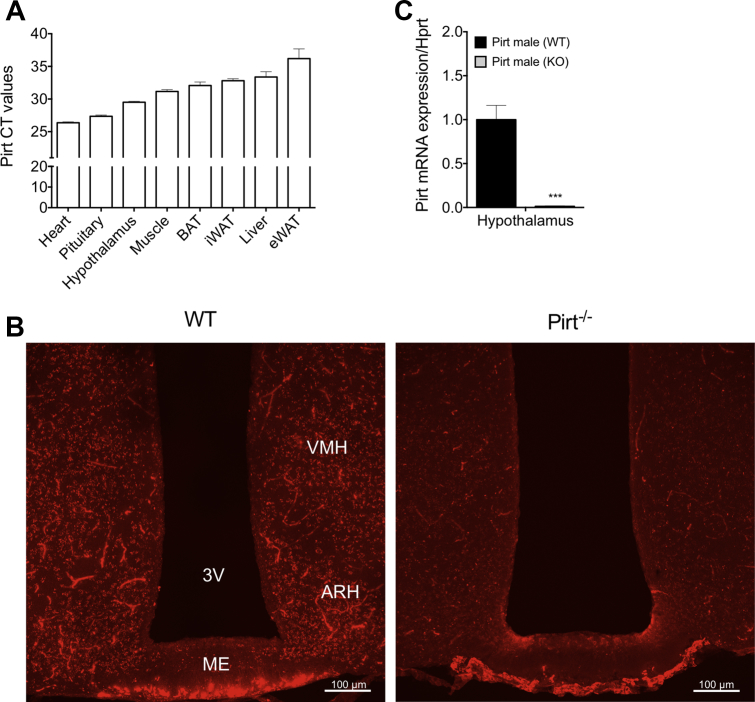
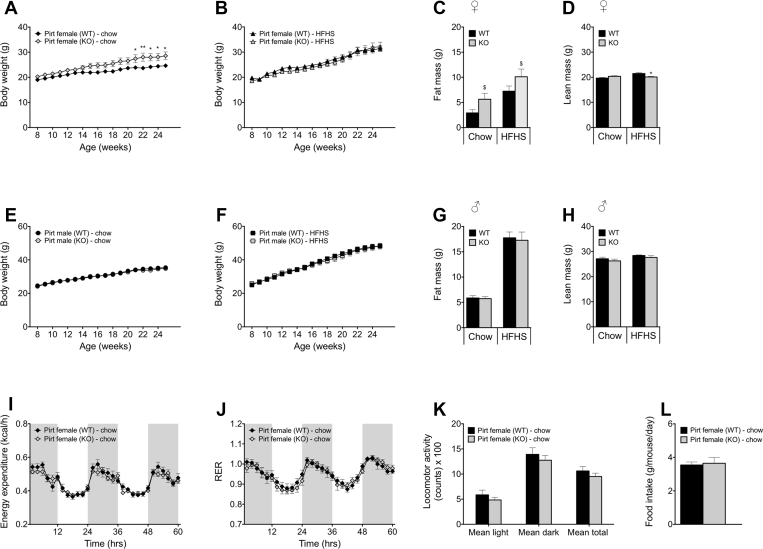
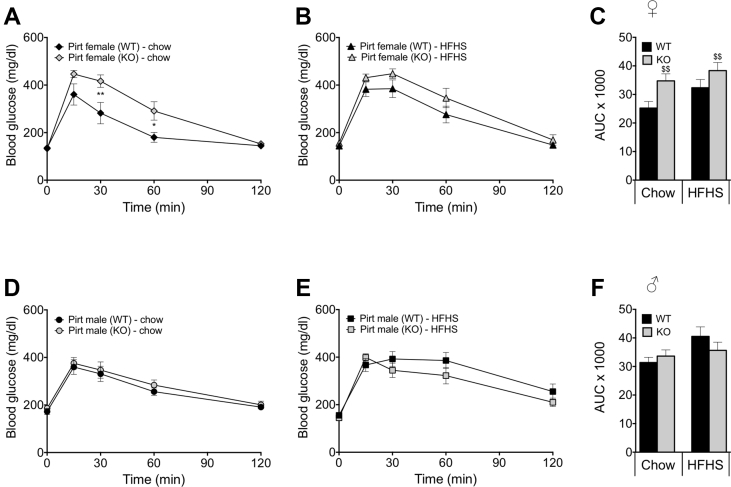
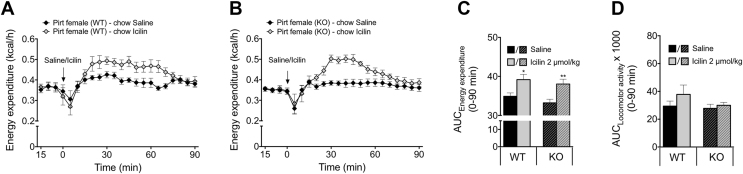
References
-
- van Marken Lichtenbelt W.D., Vanhommerig J.W., Smulders N.M., Drossaerts J.M., Kemerink G.J., Bouvy N.D. Cold-activated brown adipose tissue in healthy men. New England Journal of Medicine. 2009;360(15):1500–1508. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
