

Phototrophic extracellular electron uptake is linked to carbon dioxide fixation in the bacterium Rhodopseudomonas palustris
- PMID: 30902976
- PMCID: PMC6430793
- DOI: 10.1038/s41467-019-09377-6
Phototrophic extracellular electron uptake is linked to carbon dioxide fixation in the bacterium Rhodopseudomonas palustris
Abstract
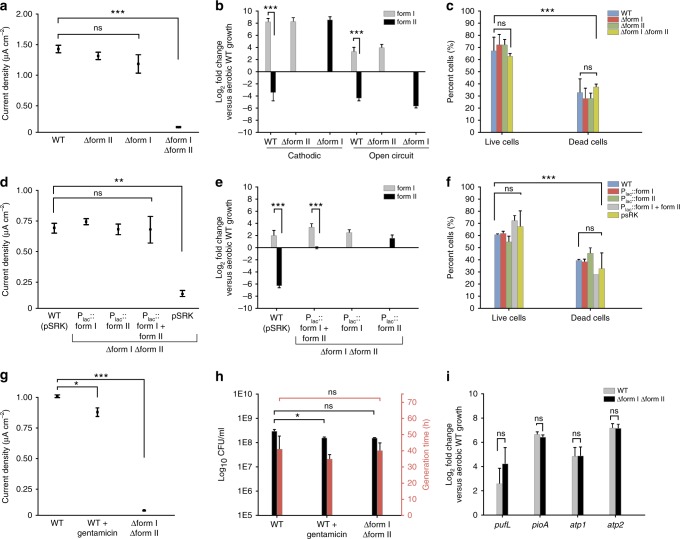
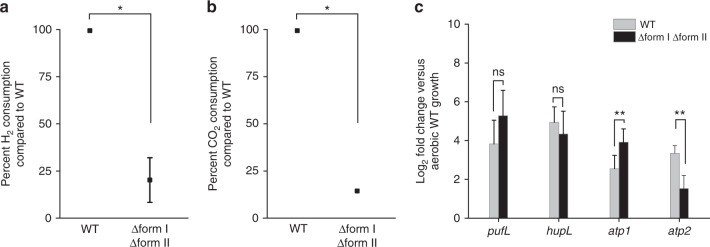
Extracellular electron uptake (EEU) is the ability of microbes to take up electrons from solid-phase conductive substances such as metal oxides. EEU is performed by prevalent phototrophic bacterial genera, but the electron transfer pathways and the physiological electron sinks are poorly understood. Here we show that electrons enter the photosynthetic electron transport chain during EEU in the phototrophic bacterium Rhodopseudomonas palustris TIE-1. Cathodic electron flow is also correlated with a highly reducing intracellular redox environment. We show that reducing equivalents are used for carbon dioxide (CO2) fixation, which is the primary electron sink. Deletion of the genes encoding ruBisCO (the CO2-fixing enzyme of the Calvin-Benson-Bassham cycle) leads to a 90% reduction in EEU. This work shows that phototrophs can directly use solid-phase conductive substances for electron transfer, energy transduction, and CO2 fixation.
Conflict of interest statement
The authors declare no competing interests.
Figures





References
-
- Kirchman, D. L. in Processes in Microbial Ecology 55–78 (Oxford University Press, Oxford, 2018).
-
- Imhoff J. F. Anoxygenic Photosynthetic Bacteria (Blankenship, R. E., Madigan, M. T. & Bauer, C. E. eds) 1–15 (Kluwer Academic Publishers, 1995).
-
- Tremblay PL, Angenent LT, Zhang T. Extracellular electron uptake: among autotrophs and mediated by surfaces. Trends Biotechnol. 2016;35:1–12. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
