

Mitochondrial Substrate-Level Phosphorylation as Energy Source for Glioblastoma: Review and Hypothesis
- PMID: 30909720
- PMCID: PMC6311572
- DOI: 10.1177/1759091418818261
Mitochondrial Substrate-Level Phosphorylation as Energy Source for Glioblastoma: Review and Hypothesis
Abstract
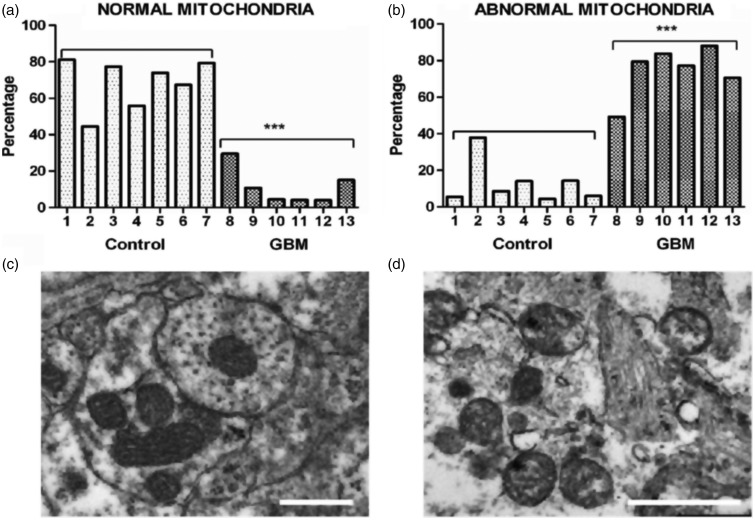
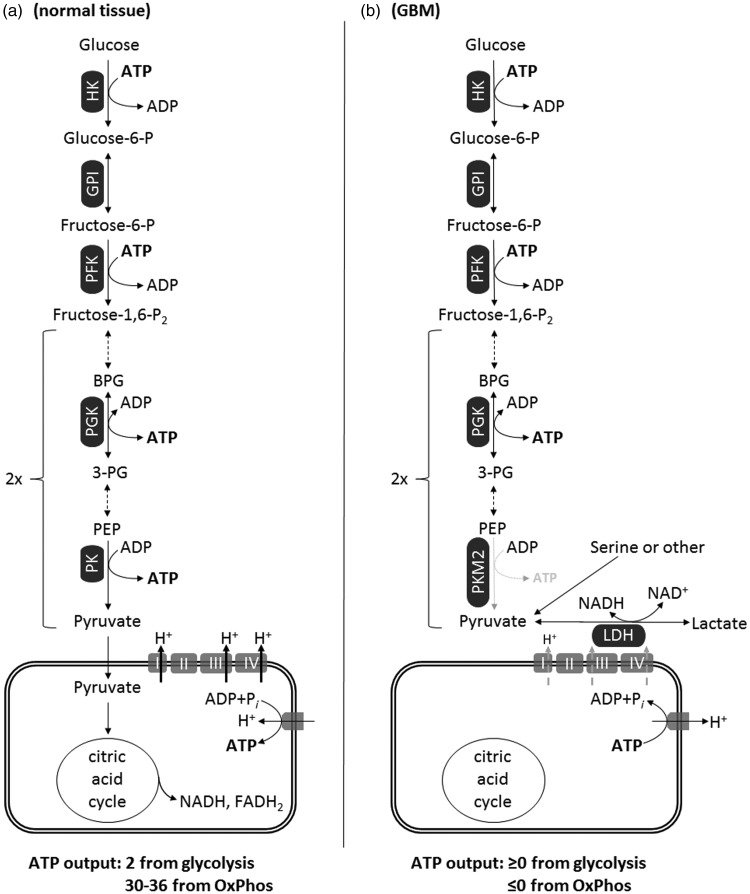
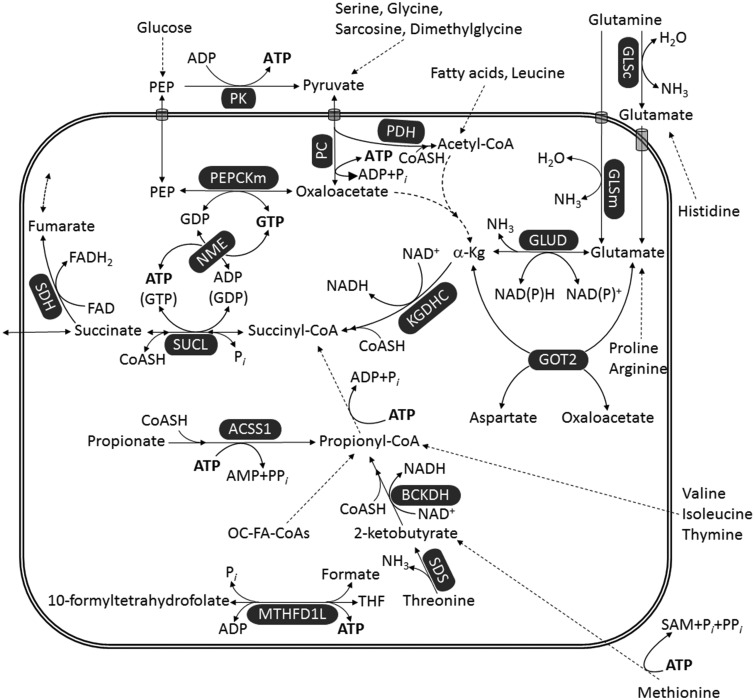
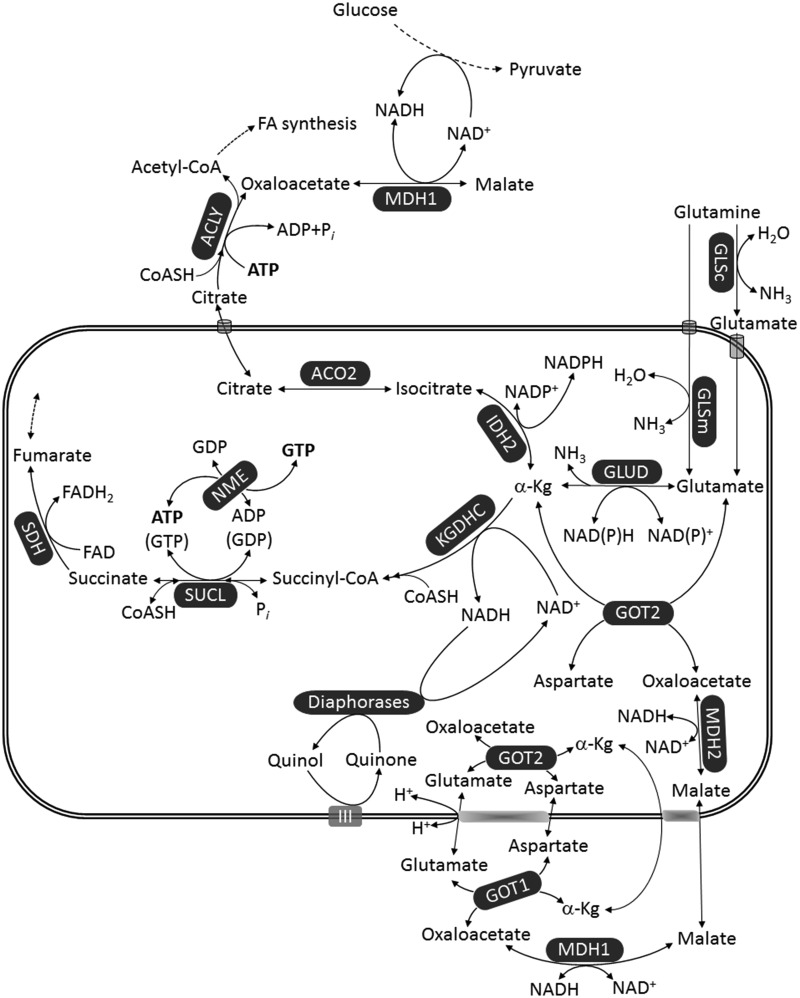
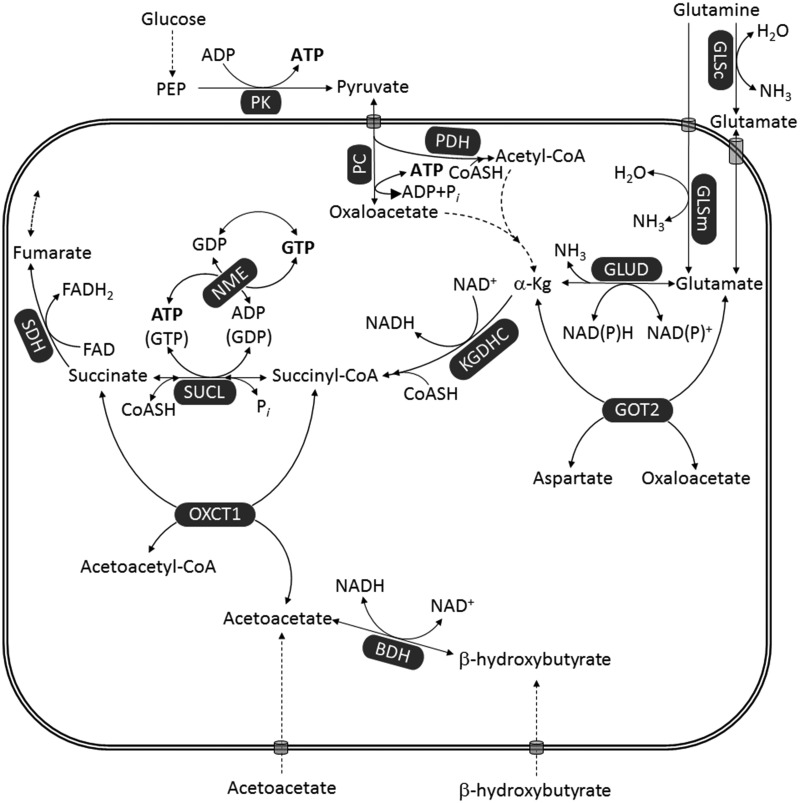
Glioblastoma multiforme (GBM) is the most common and malignant of the primary adult brain cancers. Ultrastructural and biochemical evidence shows that GBM cells exhibit mitochondrial abnormalities incompatible with energy production through oxidative phosphorylation (OxPhos). Under such conditions, the mitochondrial F0-F1 ATP synthase operates in reverse at the expense of ATP hydrolysis to maintain a moderate membrane potential. Moreover, expression of the dimeric M2 isoform of pyruvate kinase in GBM results in diminished ATP output, precluding a significant ATP production from glycolysis. If ATP synthesis through both glycolysis and OxPhos was impeded, then where would GBM cells obtain high-energy phosphates for growth and invasion? Literature is reviewed suggesting that the succinate-CoA ligase reaction in the tricarboxylic acid cycle can substantiate sufficient ATP through mitochondrial substrate-level phosphorylation (mSLP) to maintain GBM growth when OxPhos is impaired. Production of high-energy phosphates would be supported by glutaminolysis-a hallmark of GBM metabolism-through the sequential conversion of glutamine → glutamate → alpha-ketoglutarate → succinyl CoA → succinate. Equally important, provision of ATP through mSLP would maintain the adenine nucleotide translocase in forward mode, thus preventing the reverse-operating F0-F1 ATP synthase from depleting cytosolic ATP reserves. Because glucose and glutamine are the primary fuels driving the rapid growth of GBM and most tumors for that matter, simultaneous restriction of these two substrates or inhibition of mSLP should diminish cancer viability, growth, and invasion.
Keywords: Warburg; bioenergetics; gliomas; therapies.
Figures
References
-
- Ahluwalia G. S., Grem J. L., Hao Z., Cooney D. A. (1990). Metabolism and action of amino acid analog anti-cancer agents. Pharmacol Ther, 46, 243–271. - PubMed
-
- Ahmadloo N., Kani A. A., Mohammadianpanah M., Nasrolahi H., Omidvari S., Mosalaei A., Ansari M. (2013). Treatment outcome and prognostic factors of adult glioblastoma multiforme. J Egypt Natl Canc Inst, 25, 21–30. - PubMed
-
- Alam N. A., et al. (2003). Genetic and functional analyses of FH mutations in multiple cutaneous and uterine leiomyomatosis, hereditary leiomyomatosis and renal cancer, and fumarate hydratase deficiency. Hum Mol Genet, 12, 1241–1252. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Medical
