

Skin Wounding-Induced Monocyte Expansion in Mice Is Not Abrogated by IL-1 Receptor 1 Deficiency
- PMID: 30910860
- PMCID: PMC6478552
- DOI: 10.4049/jimmunol.1801481
Skin Wounding-Induced Monocyte Expansion in Mice Is Not Abrogated by IL-1 Receptor 1 Deficiency
Abstract
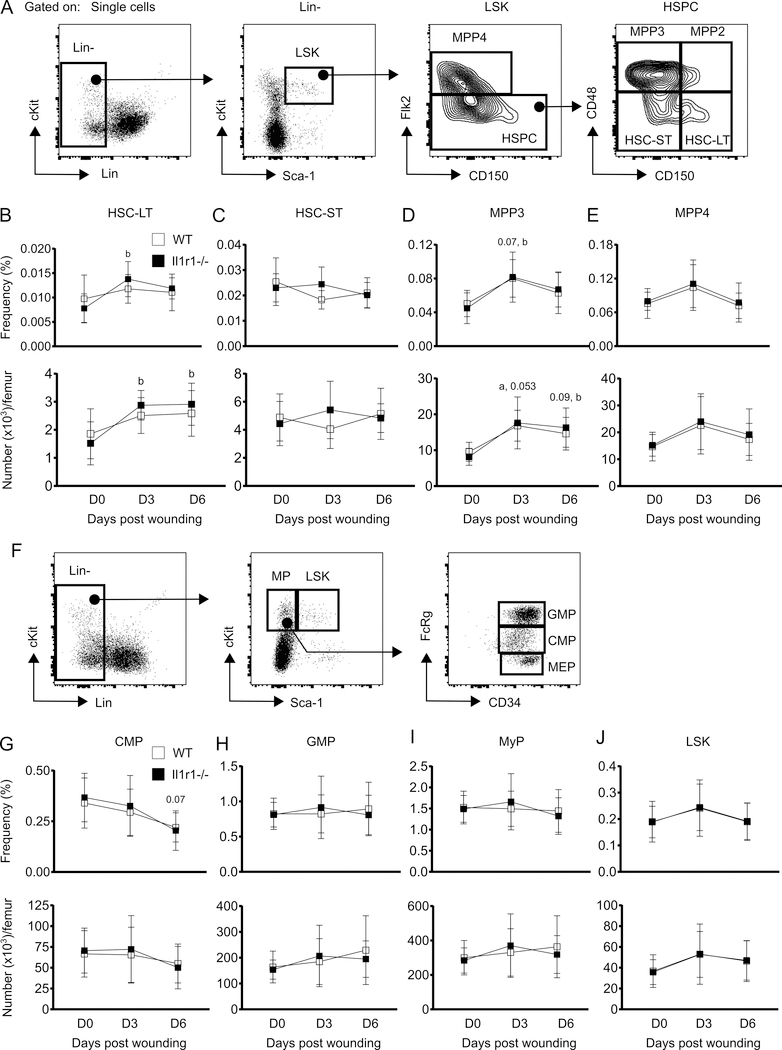
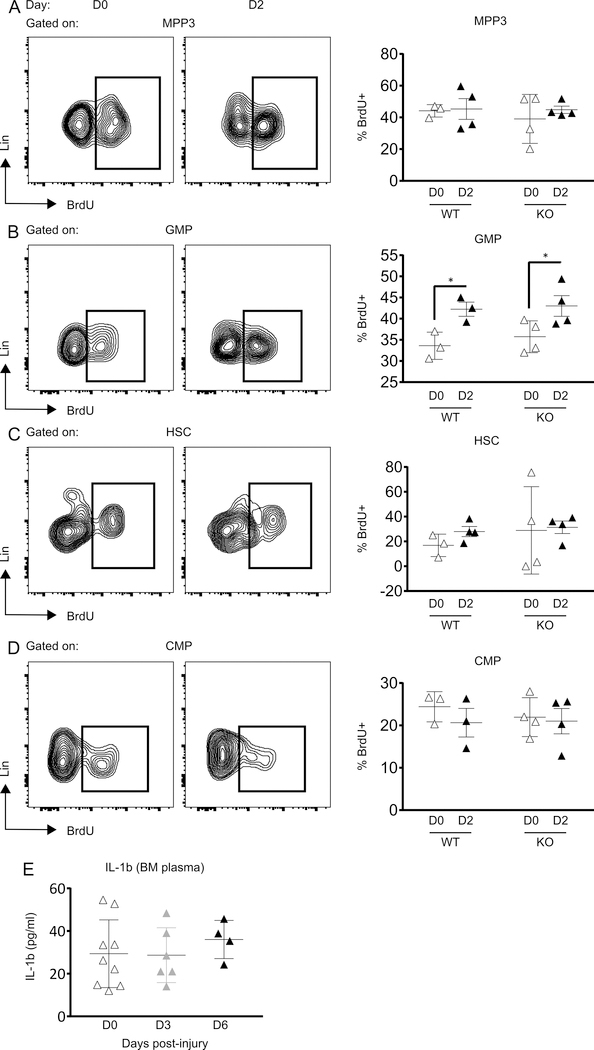
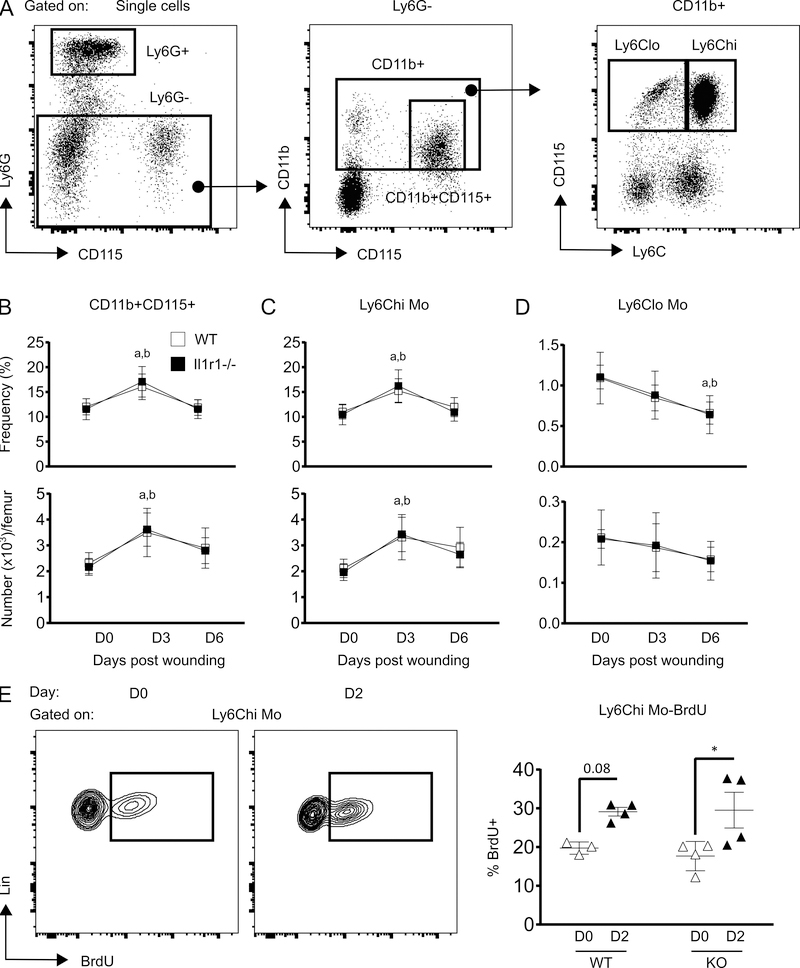
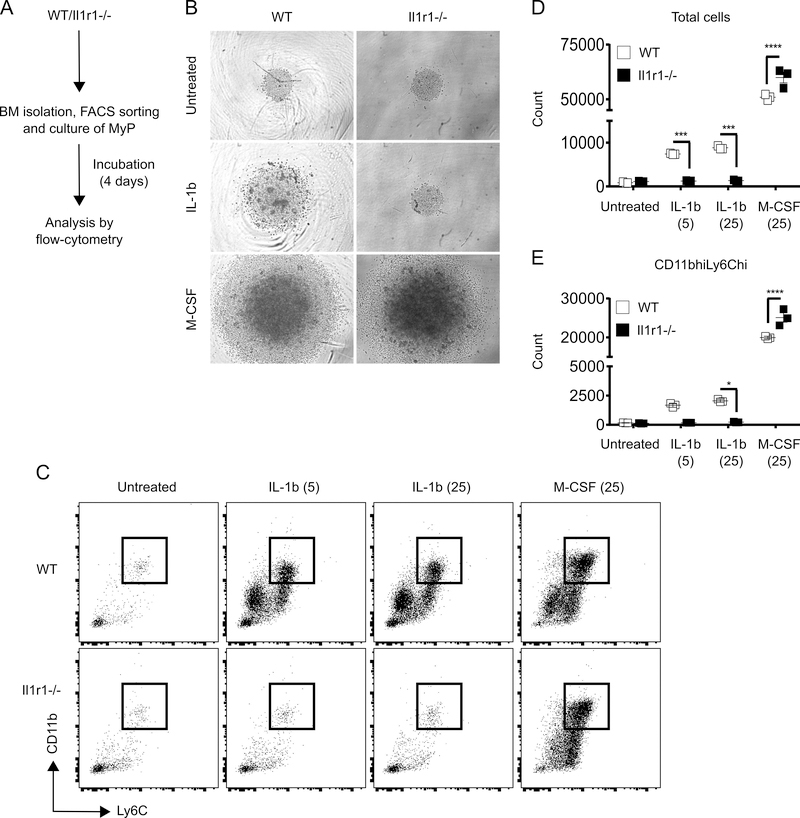
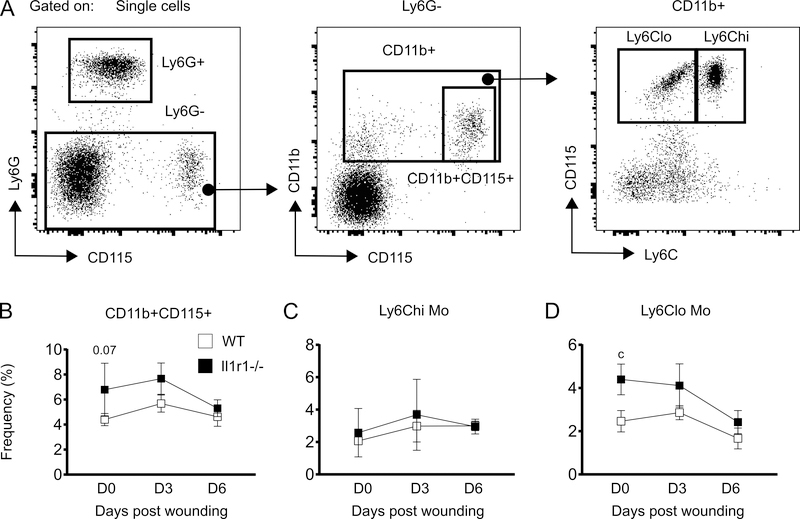
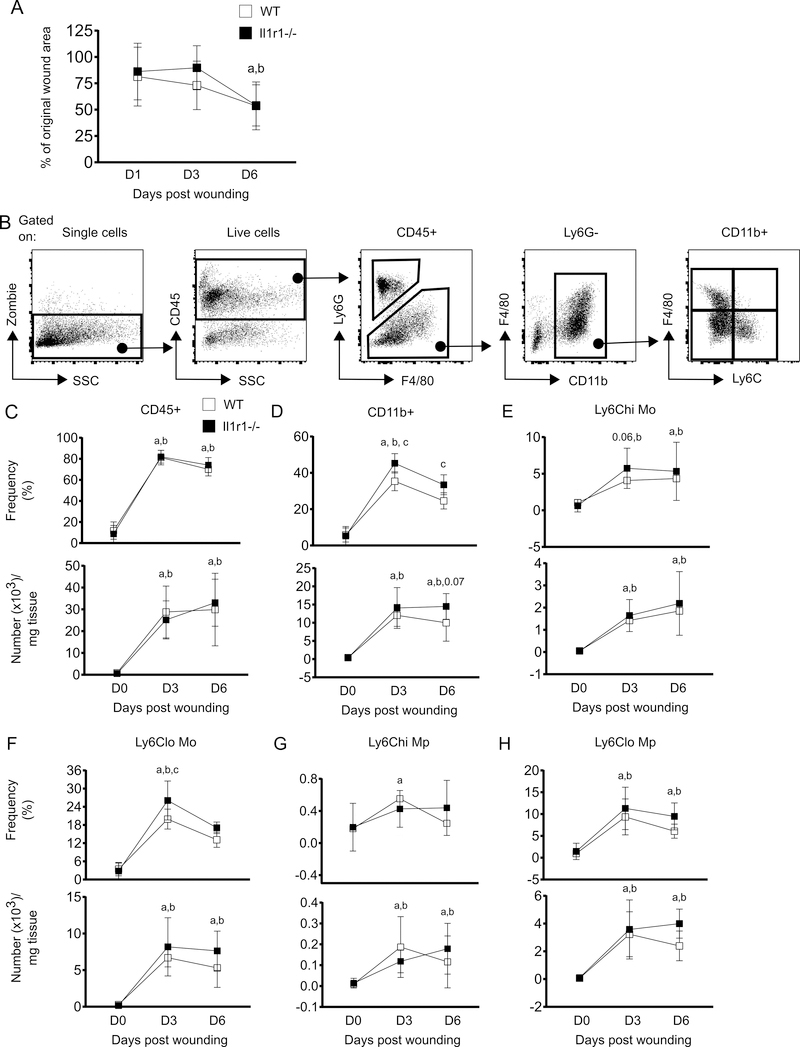
The aim of this study was to determine whether skin wounding induces monocyte (Mo) expansion in bone marrow and whether IL-1R1 signaling regulates this process. Our data show that skin wounding increases myeloid lineage-committed multipotent progenitors (MPP3 subset) and Mo in bone marrow, but this expansion is not impaired in Il1r1-/- mice. We also demonstrate that M-CSF-induced differentiation of myeloid progenitors into Mo is not impaired by the loss of IL-1R1 ex vivo, indicating that IL-R1 deficiency does not abrogate myeloid progenitor differentiation potential. In addition, we observed modestly delayed wound closure in Il1r1-/- mice associated with higher frequency of Ly6Clo Mo in the circulation at baseline and in wounds early after injury. Thus, in contrast to other models of inflammation that involve IL-1R1-dependent monopoiesis, our results demonstrate that skin wounding induces Mo progenitor and Mo expansion independently of IL-1R1 signaling.
Copyright © 2019 by The American Association of Immunologists, Inc.
Figures
References
-
- Lucas T, Waisman A, Ranjan R, Roes J, Krieg T, Muller W, Roers A, and Eming SA 2010. Differential roles of macrophages in diverse phases of skin repair. J Immunol 184: 3964–3977. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
