

Mutations in the PKM2 exon-10 region are associated with reduced allostery and increased nuclear translocation
- PMID: 30911680
- PMCID: PMC6420622
- DOI: 10.1038/s42003-019-0343-4
Mutations in the PKM2 exon-10 region are associated with reduced allostery and increased nuclear translocation
Abstract
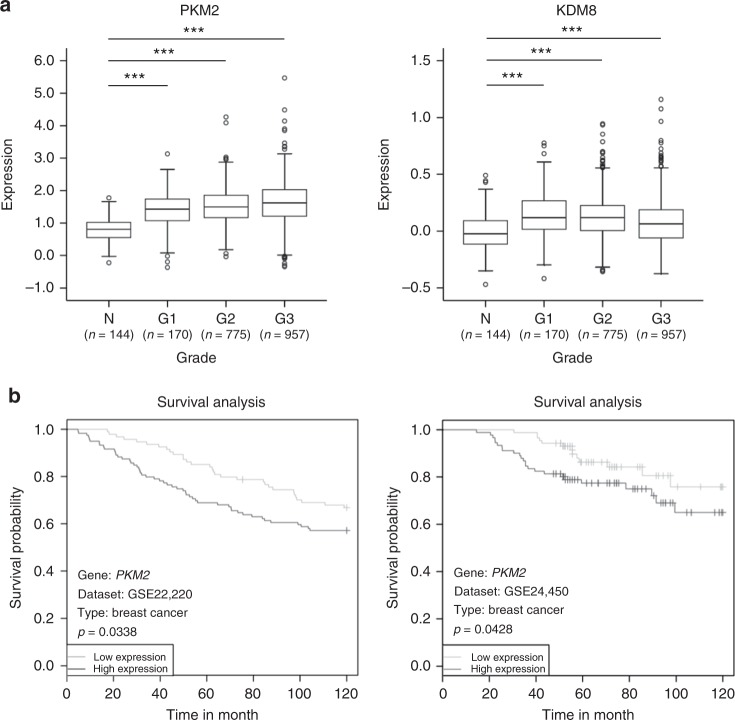
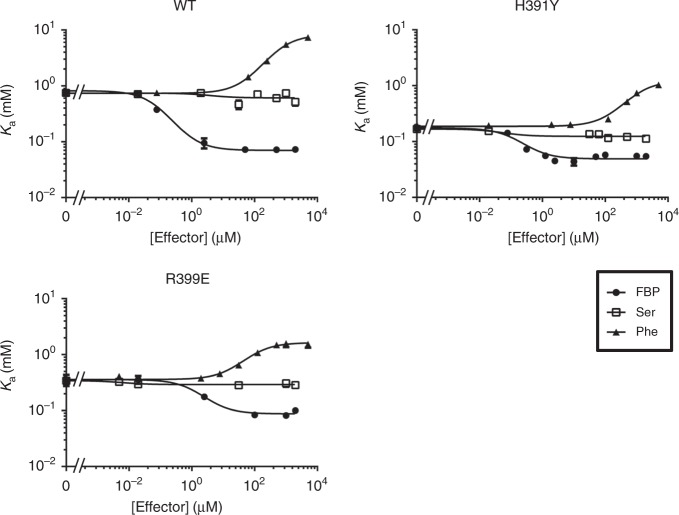
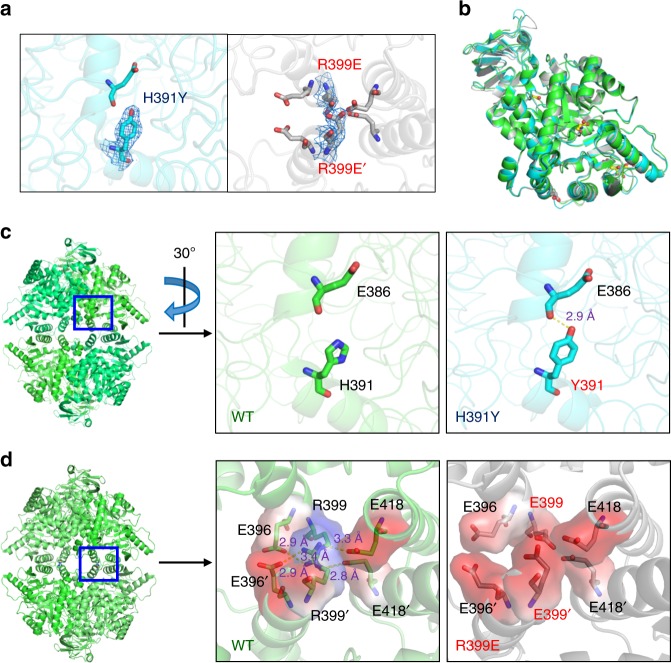
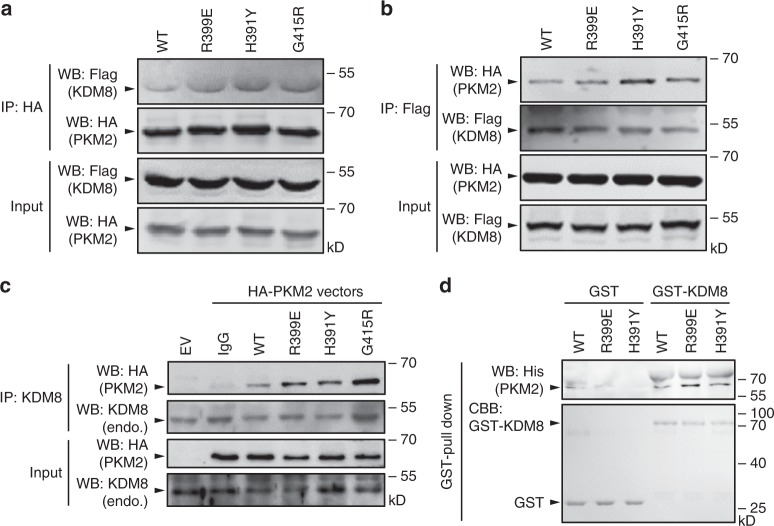
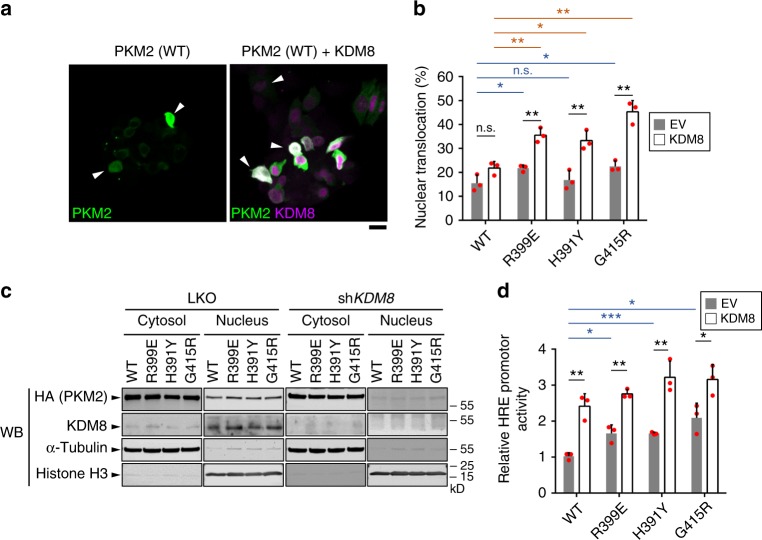
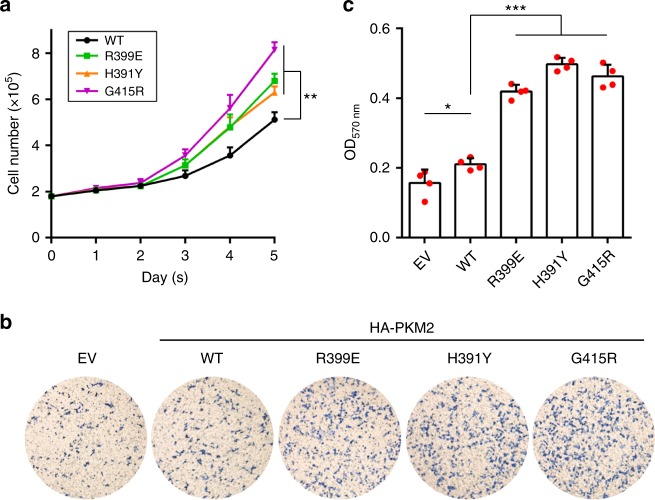
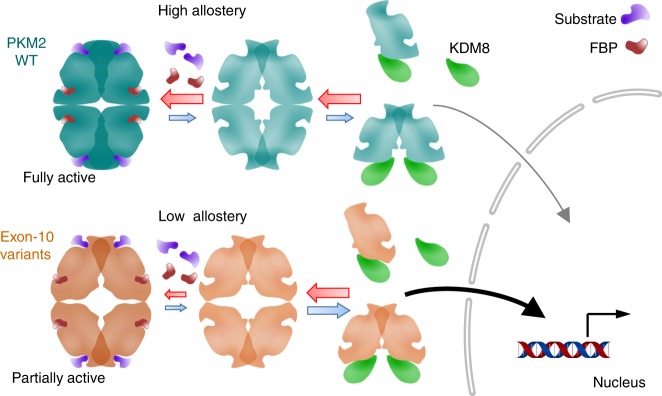
PKM2 is a key metabolic enzyme central to glucose metabolism and energy expenditure. Multiple stimuli regulate PKM2's activity through allosteric modulation and post-translational modifications. Furthermore, PKM2 can partner with KDM8, an oncogenic demethylase and enter the nucleus to serve as a HIF1α co-activator. Yet, the mechanistic basis of the exon-10 region in allosteric regulation and nuclear translocation remains unclear. Here, we determined the crystal structures and kinetic coupling constants of exon-10 tumor-related mutants (H391Y and R399E), showing altered structural plasticity and reduced allostery. Immunoprecipitation analysis revealed increased interaction with KDM8 for H391Y, R399E, and G415R. We also found a higher degree of HIF1α-mediated transactivation activity, particularly in the presence of KDM8. Furthermore, overexpression of PKM2 mutants significantly elevated cell growth and migration. Together, PKM2 exon-10 mutations lead to structure-allostery alterations and increased nuclear functions mediated by KDM8 in breast cancer cells. Targeting the PKM2-KDM8 complex may provide a potential therapeutic intervention.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Garnett ME, Dyson RD, Dost FN. Pyruvate kinase isozyme changes in parenchymal cells of regenerating rat liver. J. Biol. Chem. 1974;249:5222–5226. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
