

Crystal structure and activity-based labeling reveal the mechanisms for linkage-specific substrate recognition by deubiquitinase USP9X
- PMID: 30914461
- PMCID: PMC6462090
- DOI: 10.1073/pnas.1815027116
Crystal structure and activity-based labeling reveal the mechanisms for linkage-specific substrate recognition by deubiquitinase USP9X
Abstract
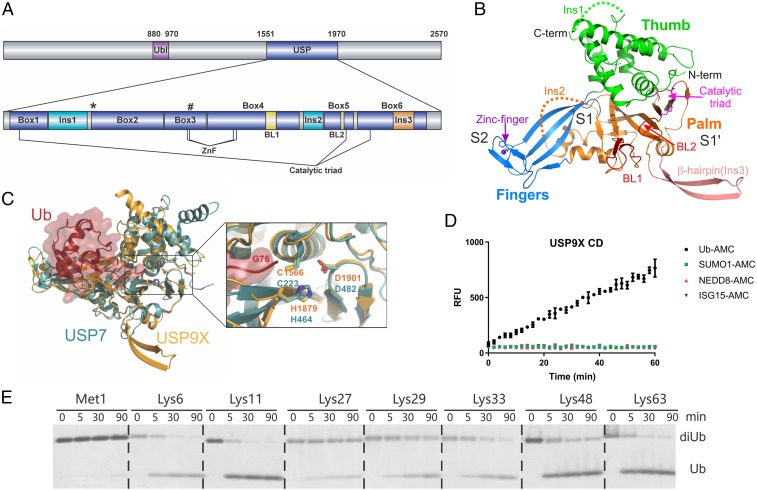
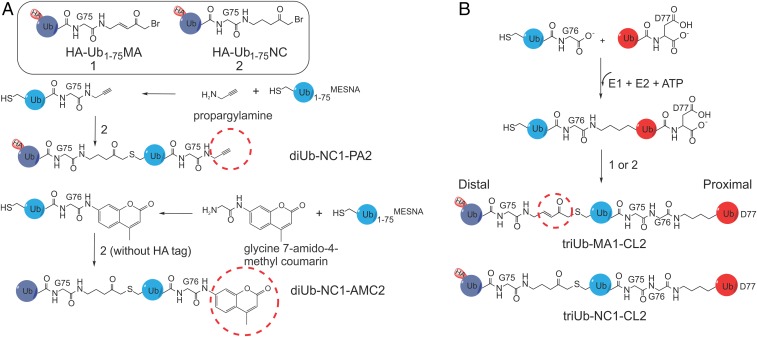
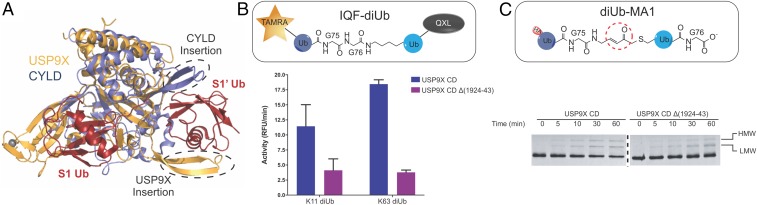
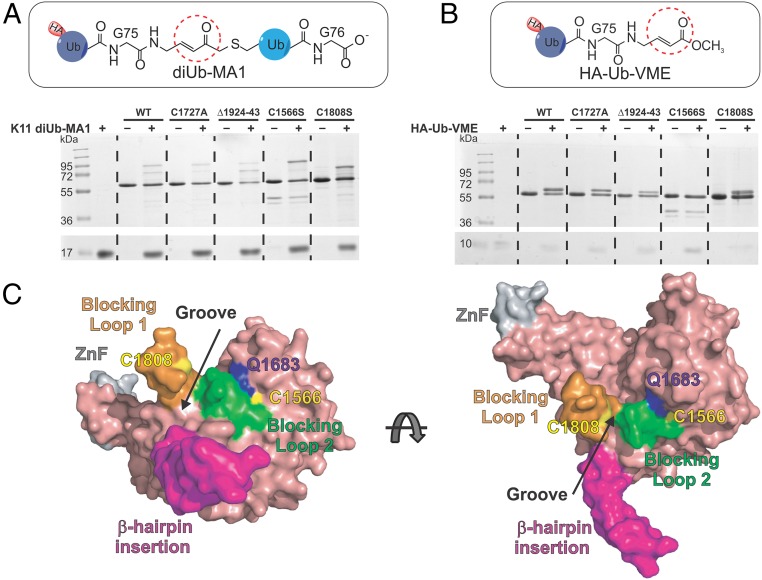
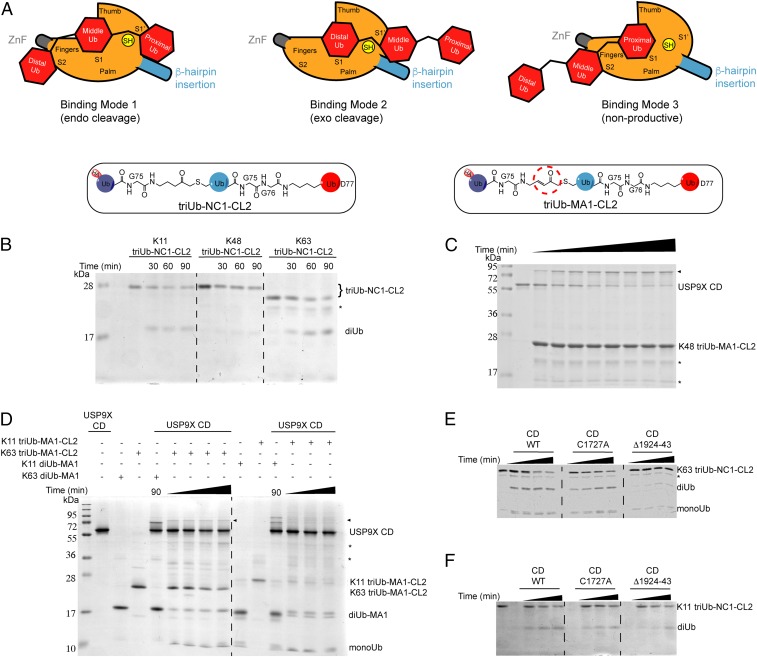
USP9X is a conserved deubiquitinase (DUB) that regulates multiple cellular processes. Dysregulation of USP9X has been linked to cancers and X-linked intellectual disability. Here, we report the crystal structure of the USP9X catalytic domain at 2.5-Å resolution. The structure reveals a canonical USP-fold comprised of fingers, palm, and thumb subdomains, as well as an unusual β-hairpin insertion. The catalytic triad of USP9X is aligned in an active configuration. USP9X is exclusively active against ubiquitin (Ub) but not Ub-like modifiers. Cleavage assays with di-, tri-, and tetraUb chains show that the USP9X catalytic domain has a clear preference for K11-, followed by K63-, K48-, and K6-linked polyUb chains. Using a set of activity-based diUb and triUb probes (ABPs), we demonstrate that the USP9X catalytic domain has an exo-cleavage preference for K48- and endo-cleavage preference for K11-linked polyUb chains. The structure model and biochemical data suggest that the USP9X catalytic domain harbors three Ub binding sites, and a zinc finger in the fingers subdomain and the β-hairpin insertion both play important roles in polyUb chain processing and linkage specificity. Furthermore, unexpected labeling of a secondary, noncatalytic cysteine located on a blocking loop adjacent to the catalytic site by K11-diUb ABP implicates a previously unreported mechanism of polyUb chain recognition. The structural features of USP9X revealed in our study are critical for understanding its DUB activity. The new Ub-based ABPs form a set of valuable tools to understand polyUb chain processing by the cysteine protease class of DUBs.
Keywords: USP9X; activity-based probes; deubiquitinase; linkage specificity; zinc finger.
Copyright © 2019 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Nijman SM, et al. A genomic and functional inventory of deubiquitinating enzymes. Cell. 2005;123:773–786. - PubMed
-
- Ye Y, Scheel H, Hofmann K, Komander D. Dissection of USP catalytic domains reveals five common insertion points. Mol Biosyst. 2009;5:1797–1808. - PubMed
-
- Komander D, Clague MJ, Urbé S. Breaking the chains: Structure and function of the deubiquitinases. Nat Rev Mol Cell Biol. 2009;10:550–563. - PubMed
-
- Faesen AC, et al. The differential modulation of USP activity by internal regulatory domains, interactors and eight ubiquitin chain types. Chem Biol. 2011;18:1550–1561. - PubMed
-
- Hu M, et al. Crystal structure of a UBP-family deubiquitinating enzyme in isolation and in complex with ubiquitin aldehyde. Cell. 2002;111:1041–1054. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
