

Introgression of a synthetic sex ratio distortion system from Anopheles gambiae into Anopheles arabiensis
- PMID: 30914785
- PMCID: PMC6435806
- DOI: 10.1038/s41598-019-41646-8
Introgression of a synthetic sex ratio distortion system from Anopheles gambiae into Anopheles arabiensis
Erratum in
-
Publisher Correction: Introgression of a synthetic sex ratio distortion system from Anopheles gambiae into Anopheles arabiensis.Sci Rep. 2019 May 22;9(1):7915. doi: 10.1038/s41598-019-44277-1. Sci Rep. 2019. PMID: 31114001 Free PMC article.
Abstract
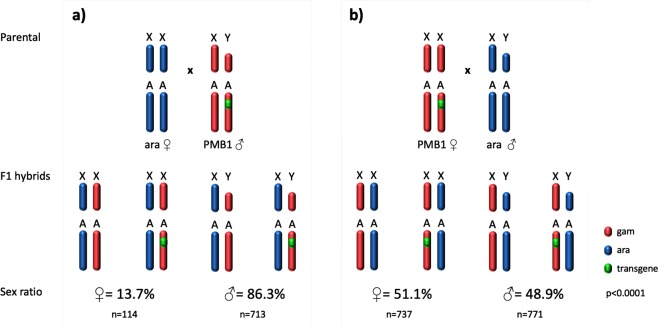
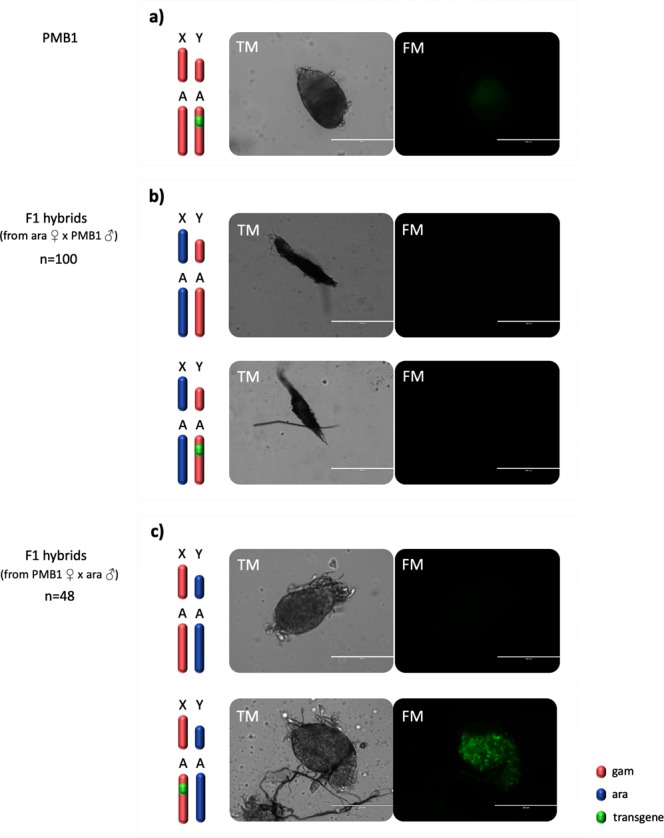
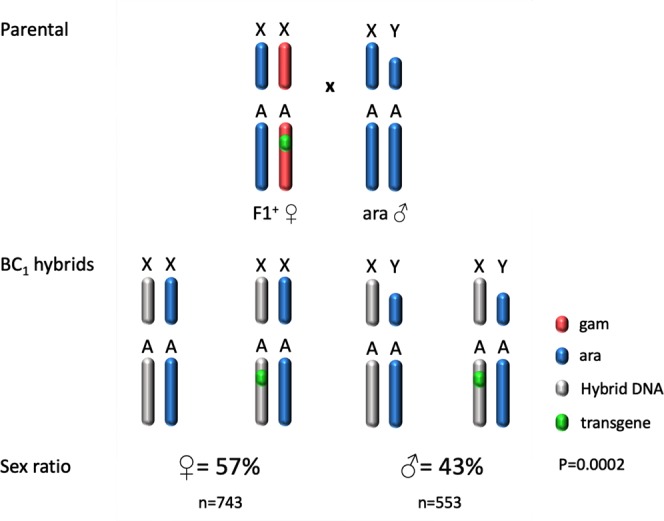
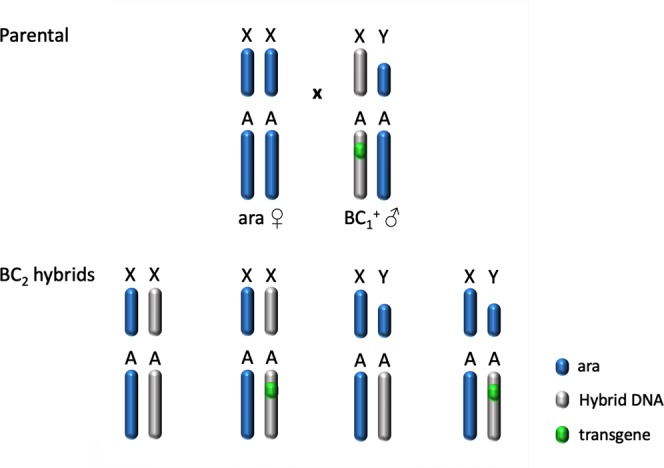
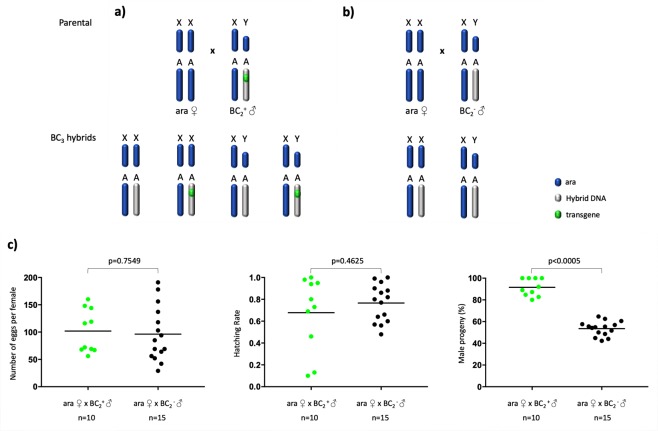
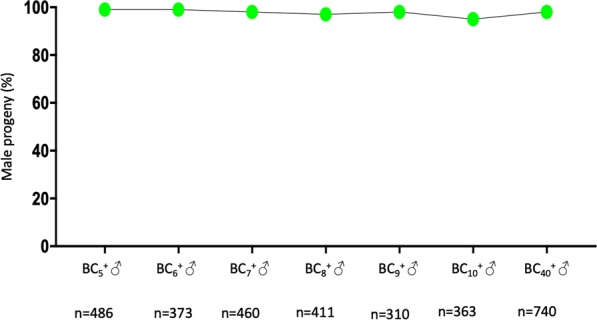
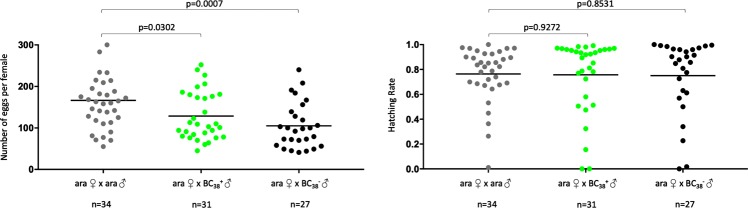
I-PpoI is a homing endonuclease that has a high cleavage activity and specificity for a conserved sequence within the ribosomal rDNA repeats, located in a single cluster on the Anopheles gambiae X chromosome. This property has been exploited to develop a synthetic sex ratio distortion system in this mosquito species. When I-PpoI is expressed from a transgene during spermatogenesis in mosquitoes, the paternal X chromosome is shredded and only Y chromosome-bearing sperm are viable, resulting in a male-biased sex ratio of >95% in the progeny. These distorter male mosquitoes can efficiently suppress caged wild-type populations, providing a powerful tool for vector control strategies. Given that malaria mosquito vectors belong to a species complex comprising at least two major vectors, we investigated whether the sex distorter I-PpoI, originally integrated in the A. gambiae genome, could be transferred via introgression to the sibling vector species Anopheles arabiensis. In compliance with Haldane's rule, F1 hybrid male sterility is known to occur in all intercrosses among members of the Anopheles gambiae complex. A scheme based on genetic crosses and transgene selection was used to bypass F1 hybrid male sterility and introgress the sex distorter I-PpoI into the A. arabiensis genetic background. Our data suggest that this sex distortion technique can be successfully applied to target A. arabiensis mosquitoes.
Conflict of interest statement
The authors declare no competing interests.
Figures

References
-
- WHO. World Malaria Report. (2018).
