

Epigenetic Regulation of Metabolism and Inflammation by Calorie Restriction
- PMID: 30915465
- PMCID: PMC6520046
- DOI: 10.1093/advances/nmy129
Epigenetic Regulation of Metabolism and Inflammation by Calorie Restriction
Abstract
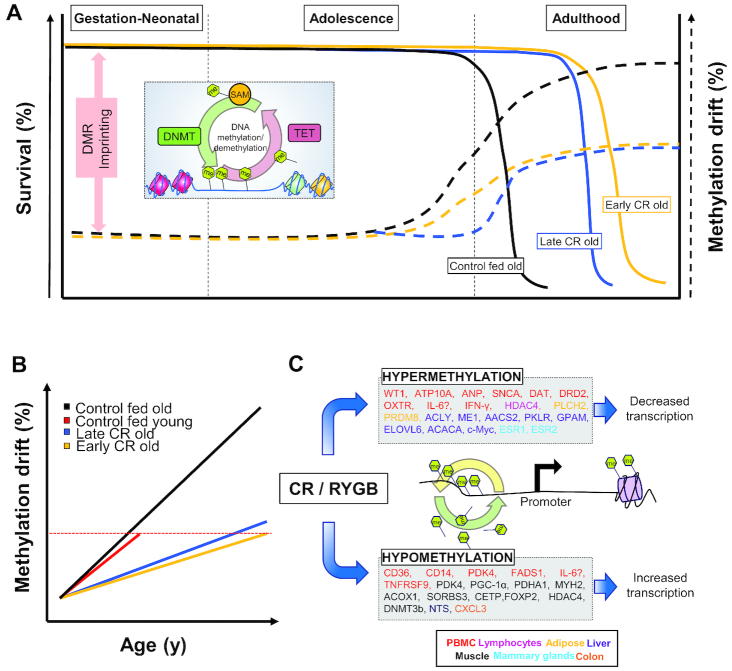
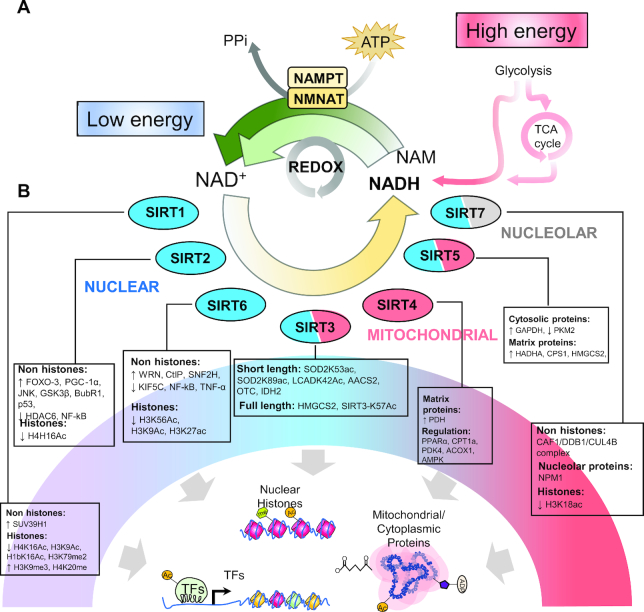
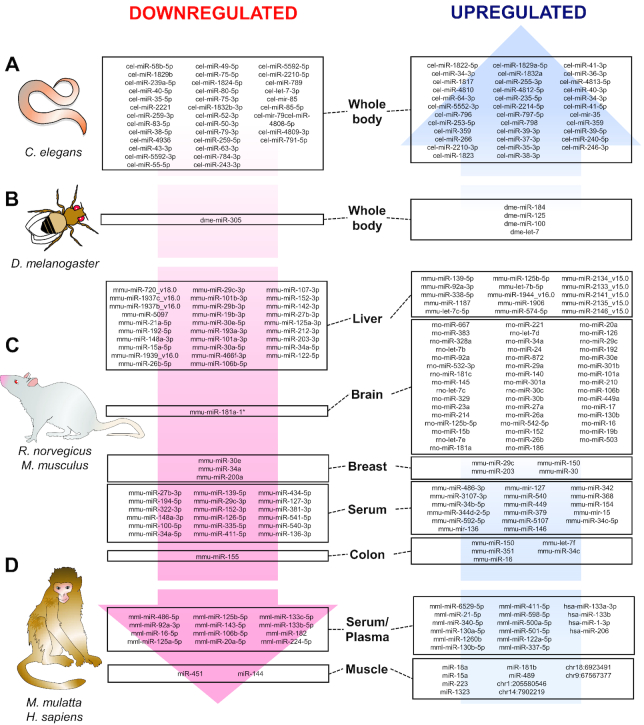
Chronic caloric restriction (CR) without malnutrition is known to affect different cellular processes such as stem cell function, cell senescence, inflammation, and metabolism. Despite the differences in the implementation of CR, the reduction of calories produces a widespread beneficial effect in noncommunicable chronic diseases, which can be explained by improvements in immuno-metabolic adaptation. Cellular adaptation that occurs in response to dietary patterns can be explained by alterations in epigenetic mechanisms such as DNA methylation, histone modifications, and microRNA. In this review, we define these modifications and systematically summarize the current evidence related to CR and the epigenome. We then explain the significance of genome-wide epigenetic modifications in the context of disease development. Although substantial evidence exists for the widespread effect of CR on longevity, there is no consensus regarding the epigenetic regulations of the underlying cellular mechanisms that lead to improved health. We provide compelling evidence that CR produces long-lasting epigenetic effects that mediate expression of genes related to immuno-metabolic processes. Epigenetic reprogramming of the underlying chronic low-grade inflammation by CR can lead to immuno-metabolic adaptations that enhance quality of life, extend lifespan, and delay chronic disease onset.
Keywords: DNA methylation; dietary restriction; energy intake; histone acetylation; microRNA; sirtuin.
Copyright © American Society for Nutrition 2019.
Figures
References
-
- Heilbronn LK, Ravussin E. Calorie restriction and aging: review of the literature and implications for studies in humans. Am J Clin Nutr. 2003;78(3):361–9. - PubMed
-
- Parra D, Gonzalez A, Martinez JA, Labayen I, Diez N. In vivo assessment of the mitochondrial response to caloric restriction in obese women by the 2-keto[1-13C]isocaproate breath test. Metabolism. 2003;52(4):463–7. - PubMed
