

Effector and Activated T Cells Induce Preterm Labor and Birth That Is Prevented by Treatment with Progesterone
- PMID: 30918041
- PMCID: PMC6570421
- DOI: 10.4049/jimmunol.1801350
Effector and Activated T Cells Induce Preterm Labor and Birth That Is Prevented by Treatment with Progesterone
Abstract
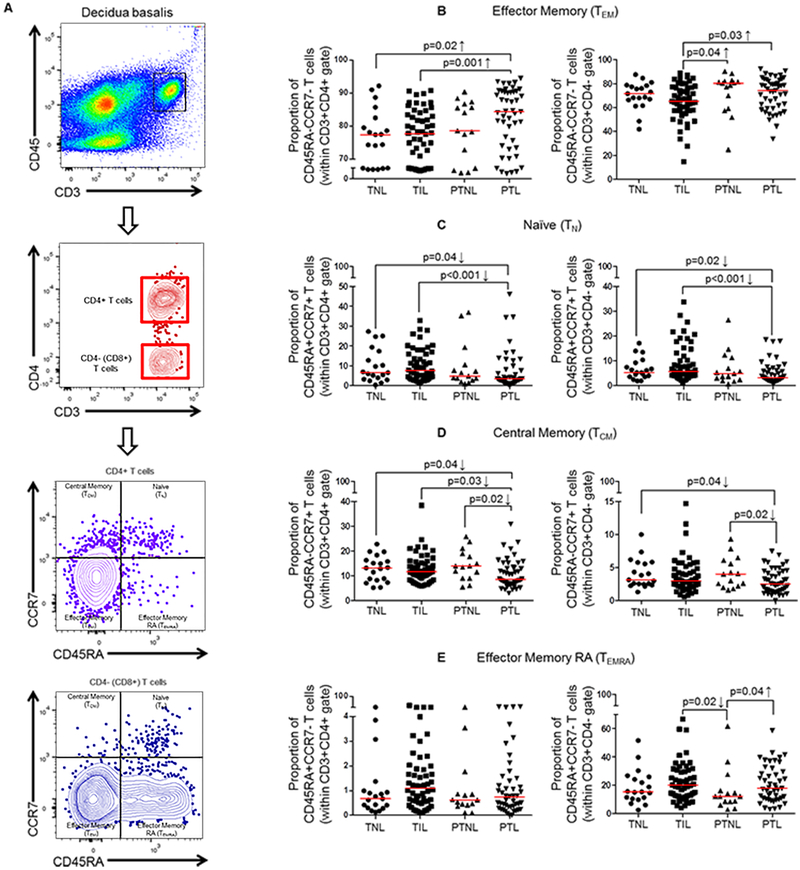
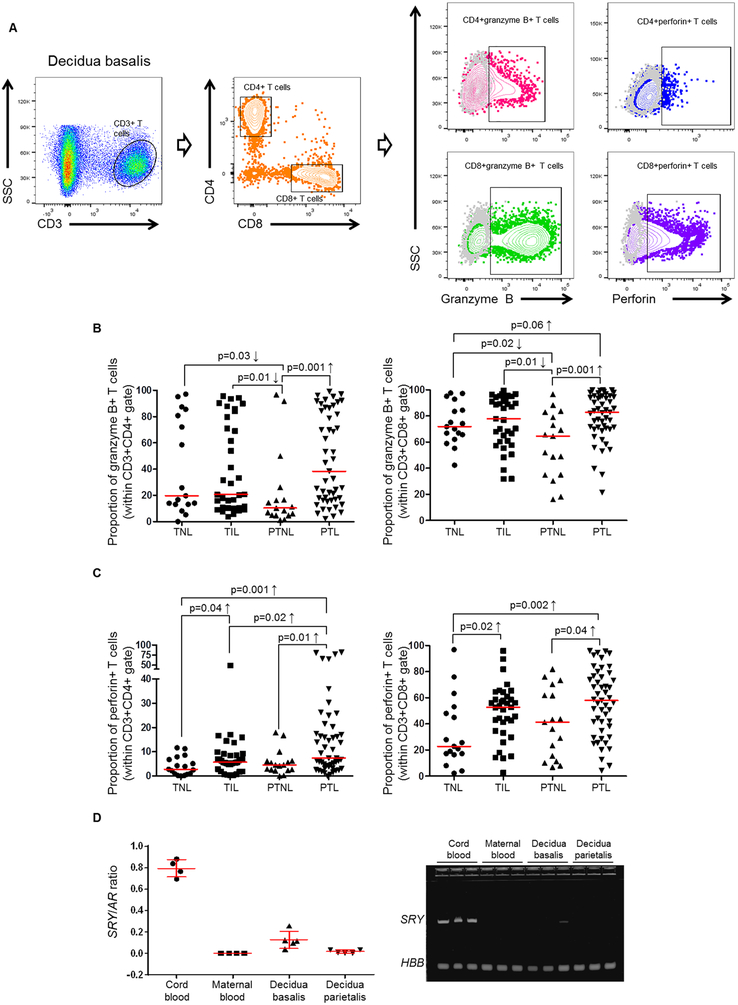
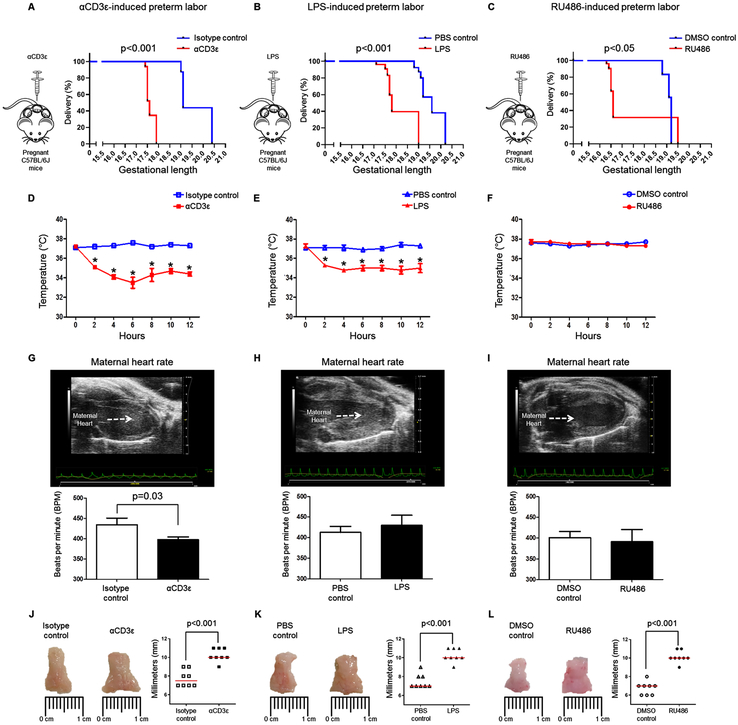
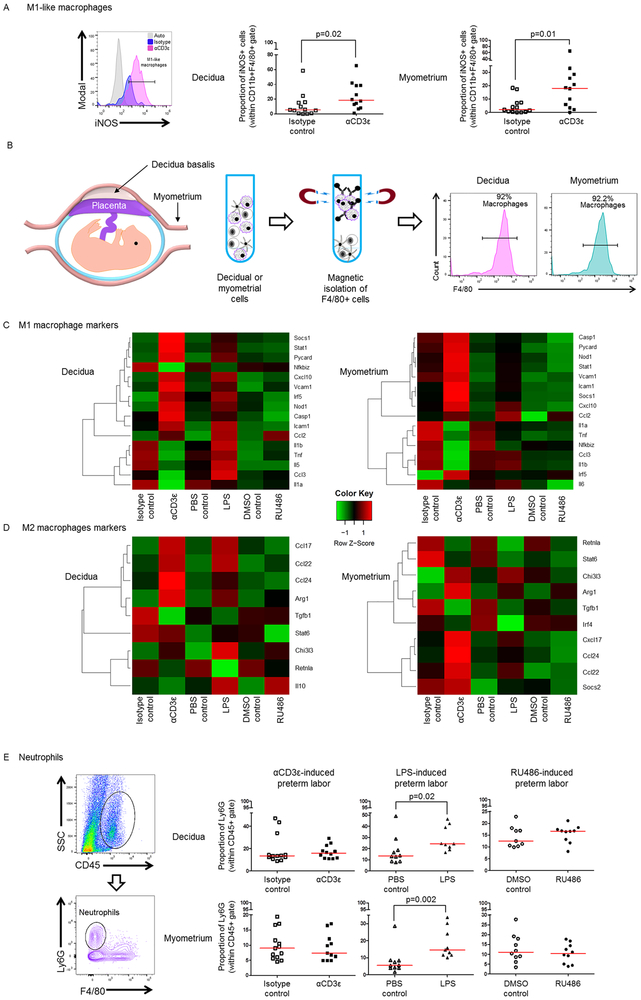
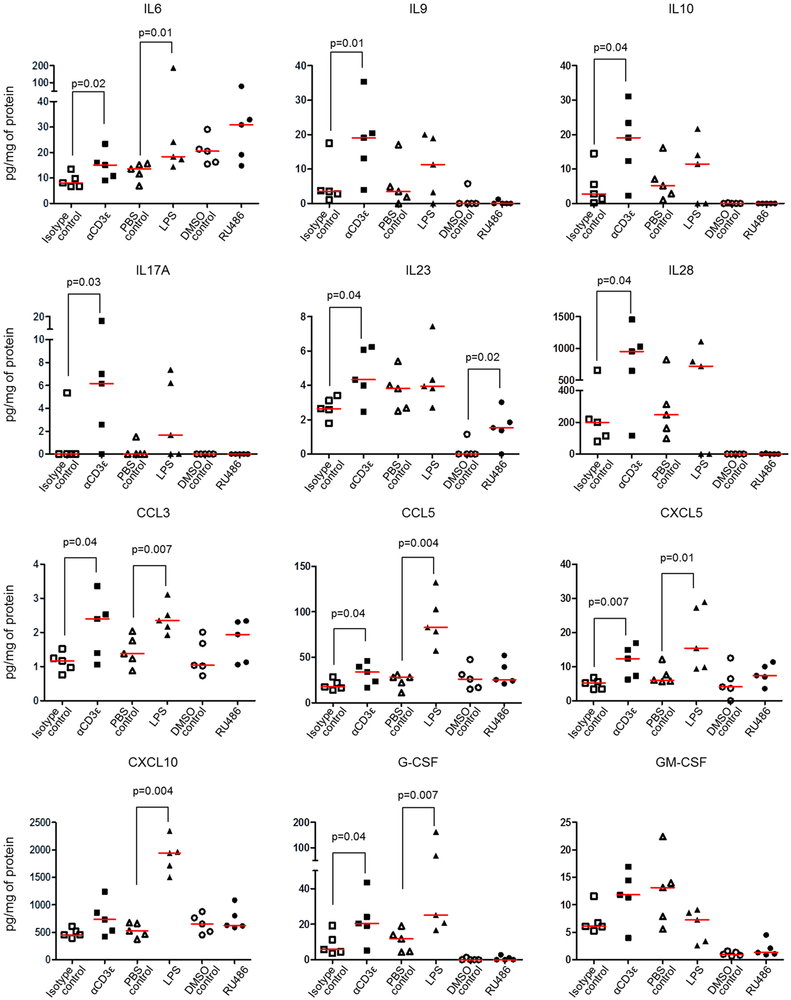
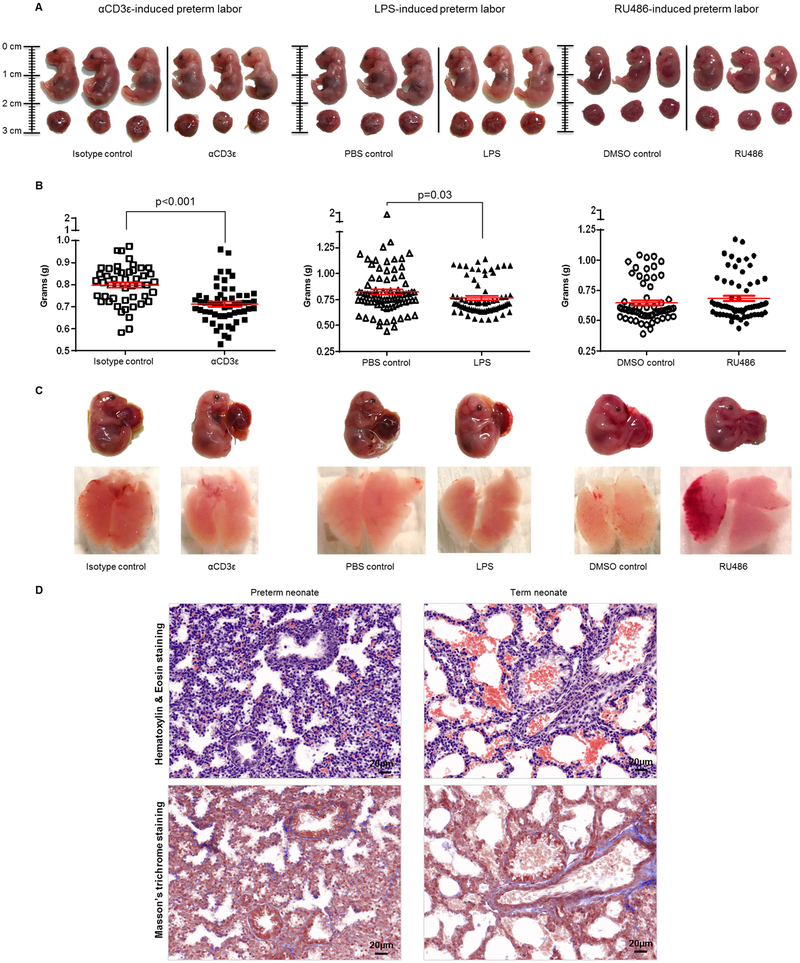
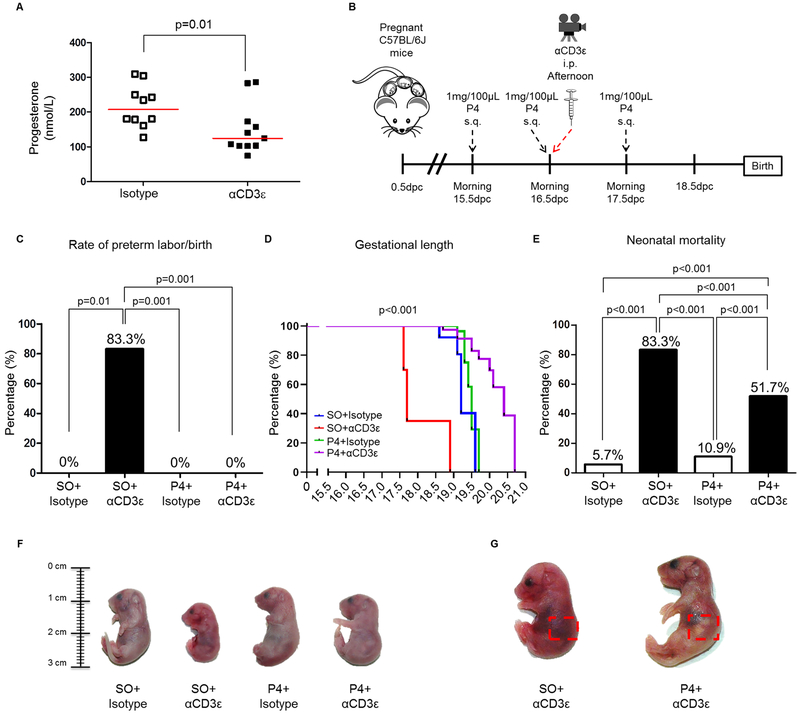
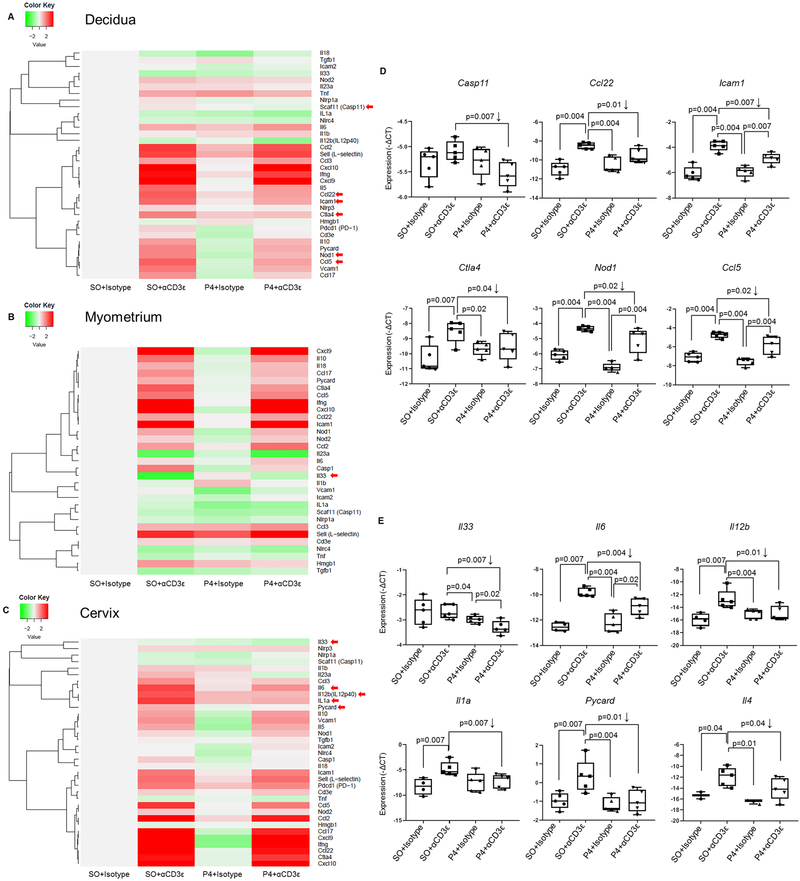
Preterm labor commonly precedes preterm birth, the leading cause of perinatal morbidity and mortality worldwide. Most research has focused on establishing a causal link between innate immune activation and pathological inflammation leading to preterm labor and birth. However, the role of maternal effector/activated T cells in the pathogenesis of preterm labor/birth is poorly understood. In this study, we first demonstrated that effector memory and activated maternal T cells expressing granzyme B and perforin are enriched at the maternal-fetal interface (decidua) of women with spontaneous preterm labor. Next, using a murine model, we reported that prior to inducing preterm birth, in vivo T cell activation caused maternal hypothermia, bradycardia, systemic inflammation, cervical dilation, intra-amniotic inflammation, and fetal growth restriction, all of which are clinical signs associated with preterm labor. In vivo T cell activation also induced B cell cytokine responses, a proinflammatory macrophage polarization, and other inflammatory responses at the maternal-fetal interface and myometrium in the absence of an increased influx of neutrophils. Finally, we showed that treatment with progesterone can serve as a strategy to prevent preterm labor/birth and adverse neonatal outcomes by attenuating the proinflammatory responses at the maternal-fetal interface and cervix induced by T cell activation. Collectively, these findings provide mechanistic evidence showing that effector and activated T cells cause pathological inflammation at the maternal-fetal interface, in the mother, and in the fetus, inducing preterm labor and birth and adverse neonatal outcomes. Such adverse effects can be prevented by treatment with progesterone, a clinically approved strategy.
Copyright © 2019 by The American Association of Immunologists, Inc.
Figures



Similar articles
-
Maternal and fetal T cells in term pregnancy and preterm labor.Cell Mol Immunol. 2020 Jul;17(7):693-704. doi: 10.1038/s41423-020-0471-2. Epub 2020 May 28. Cell Mol Immunol. 2020. PMID: 32467619 Free PMC article. Review.
-
Regulatory T Cells Play a Role in a Subset of Idiopathic Preterm Labor/Birth and Adverse Neonatal Outcomes.Cell Rep. 2020 Jul 7;32(1):107874. doi: 10.1016/j.celrep.2020.107874. Cell Rep. 2020. PMID: 32640239 Free PMC article.
-
Inhibition of the NLRP3 inflammasome can prevent sterile intra-amniotic inflammation, preterm labor/birth, and adverse neonatal outcomes†.Biol Reprod. 2019 May 1;100(5):1306-1318. doi: 10.1093/biolre/ioy264. Biol Reprod. 2019. PMID: 30596885 Free PMC article.
-
Human Chorionic Gonadotropin Has Anti-Inflammatory Effects at the Maternal-Fetal Interface and Prevents Endotoxin-Induced Preterm Birth, but Causes Dystocia and Fetal Compromise in Mice.Biol Reprod. 2016 Jun;94(6):136. doi: 10.1095/biolreprod.116.139345. Epub 2016 May 4. Biol Reprod. 2016. PMID: 27146032 Free PMC article.
-
The immunobiology of preterm labor and birth: intra-amniotic inflammation or breakdown of maternal-fetal homeostasis.Reproduction. 2022 Jun 20;164(2):R11-R45. doi: 10.1530/REP-22-0046. Reproduction. 2022. PMID: 35559791 Free PMC article. Review.
Cited by
-
Interventions for Infection and Inflammation-Induced Preterm Birth: a Preclinical Systematic Review.Reprod Sci. 2023 Feb;30(2):361-379. doi: 10.1007/s43032-022-00934-x. Epub 2022 Apr 14. Reprod Sci. 2023. PMID: 35426035 Free PMC article.
-
Exploring myometrial microenvironment changes at the single-cell level from nonpregnant to term pregnant states.Physiol Genomics. 2024 Jan 1;56(1):32-47. doi: 10.1152/physiolgenomics.00067.2023. Epub 2023 Nov 13. Physiol Genomics. 2024. PMID: 37955337 Free PMC article.
-
Human Leucocyte Antigen G and Murine Qa-2 Are Critical for Myeloid Derived Suppressor Cell Expansion and Activation and for Successful Pregnancy Outcome.Front Immunol. 2022 Jan 17;12:787468. doi: 10.3389/fimmu.2021.787468. eCollection 2021. Front Immunol. 2022. PMID: 35111157 Free PMC article.
-
The Inflammatory Cytokine Imbalance for Miscarriage, Pregnancy Loss and COVID-19 Pneumonia.Front Immunol. 2022 Mar 11;13:861245. doi: 10.3389/fimmu.2022.861245. eCollection 2022. Front Immunol. 2022. PMID: 35359975 Free PMC article. Review.
-
Immune Cells in the Placental Villi Contribute to Intra-amniotic Inflammation.Front Immunol. 2020 May 22;11:866. doi: 10.3389/fimmu.2020.00866. eCollection 2020. Front Immunol. 2020. PMID: 32528468 Free PMC article.
References
-
- Blencowe H, Cousens S, Oestergaard MZ, Chou D, Moller AB, Narwal R, Adler A, Vera Garcia C, Rohde S, Say L, and Lawn JE. 2012. National, regional, and worldwide estimates of preterm birth rates in the year 2010 with time trends since 1990 for selected countries: a systematic analysis and implications. Lancet 379: 2162–2172. - PubMed
-
- Liu L, Oza S, Hogan D, Perin J, Rudan I, Lawn JE, Cousens S, Mathers C, and Black RE. 2015. Global, regional, and national causes of child mortality in 2000–13, with projections to inform post-2015 priorities: an updated systematic analysis. Lancet 385: 430–440. - PubMed
-
- Romero R, and Lockwood CJ. 2009. Pathogenesis of Spontaneous Preterm Labor In Creasy and Resnik’s Maternal-Fetal Medicine: Principles and Practice, Sixth ed. Creasy RK, Resnik R, and Iams J, eds. Elsevier, Philadelphia: 521–543.
-
- Muglia LJ, and Katz M. 2010. The enigma of spontaneous preterm birth. N Engl J Med 362: 529–535. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
