

Diversity of KIR, HLA Class I, and Their Interactions in Seven Populations of Sub-Saharan Africans
- PMID: 30918042
- PMCID: PMC6690726
- DOI: 10.4049/jimmunol.1801586
Diversity of KIR, HLA Class I, and Their Interactions in Seven Populations of Sub-Saharan Africans
Abstract
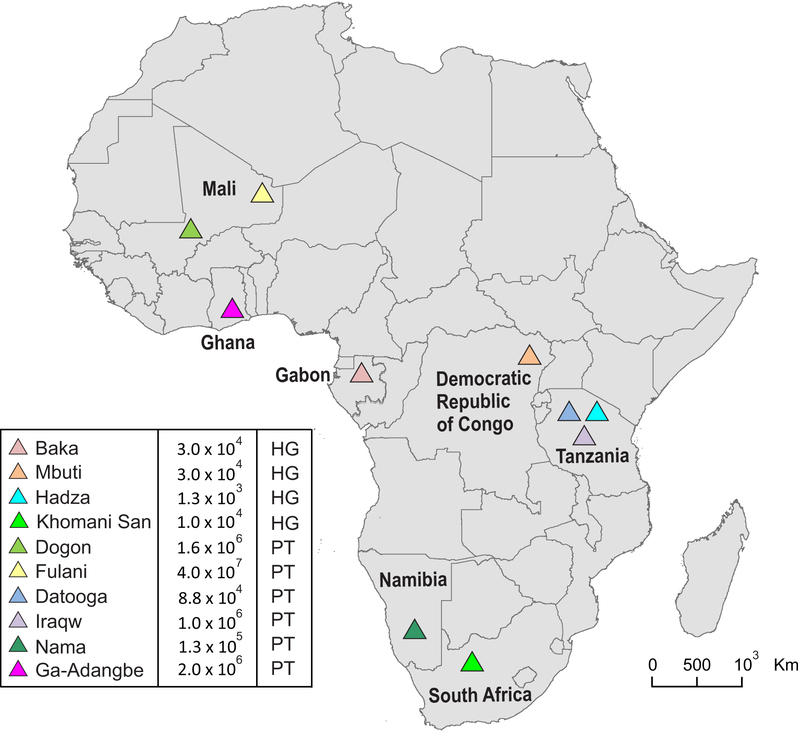
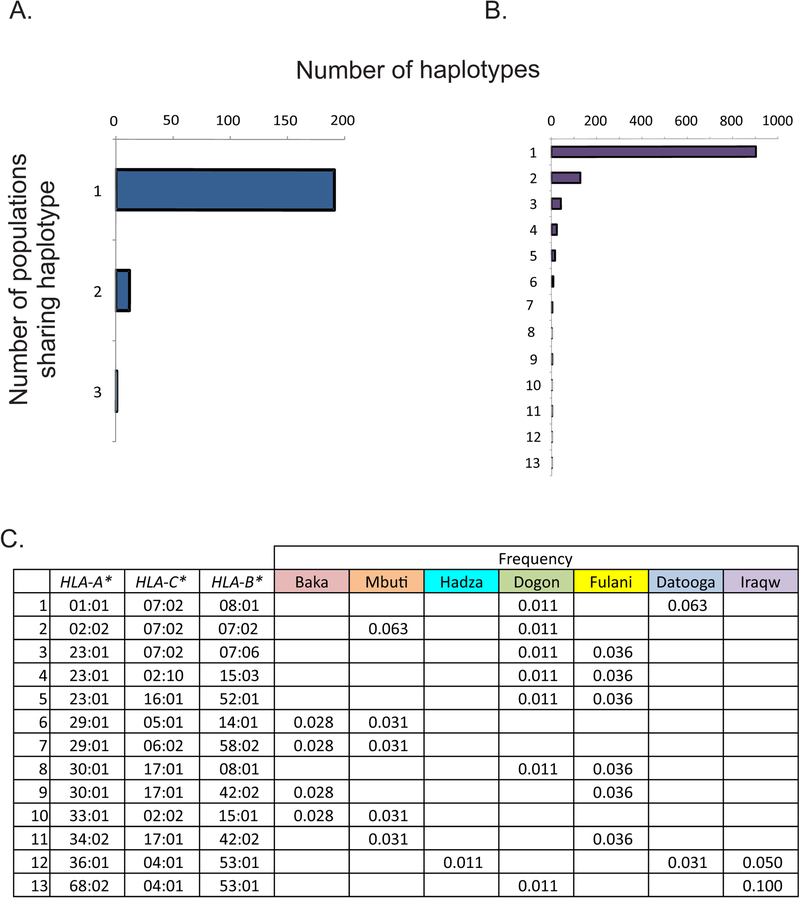
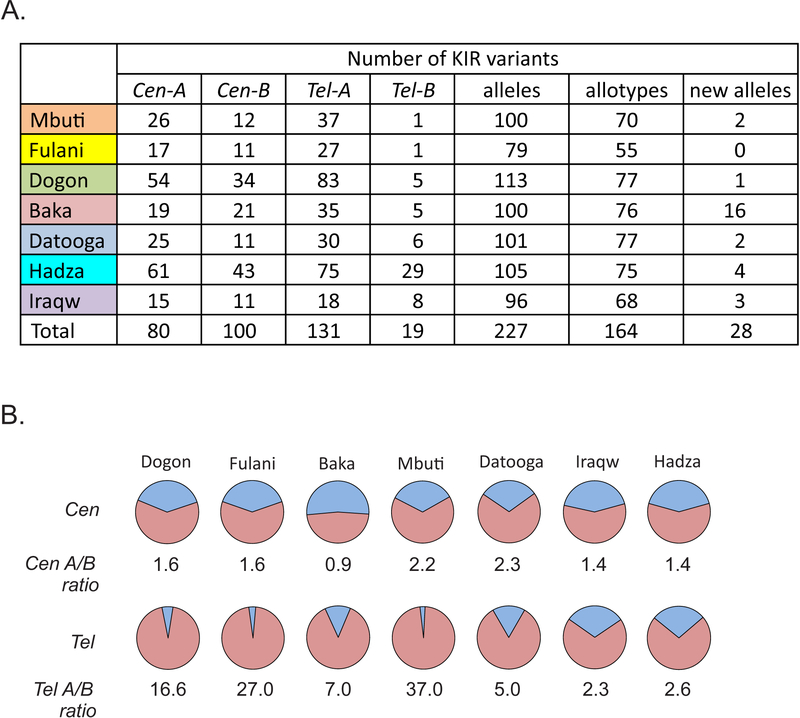
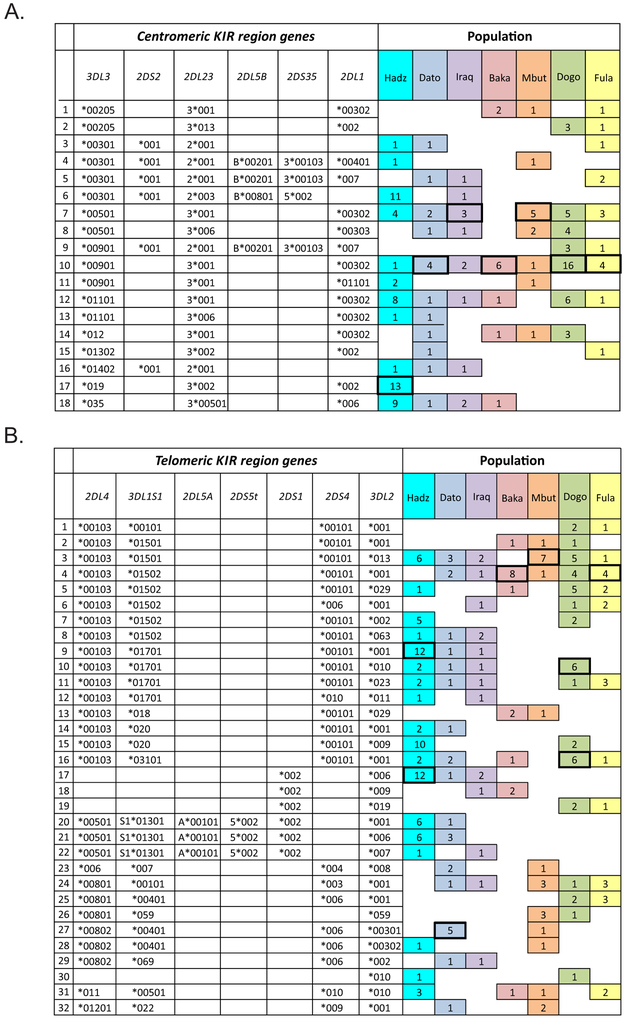
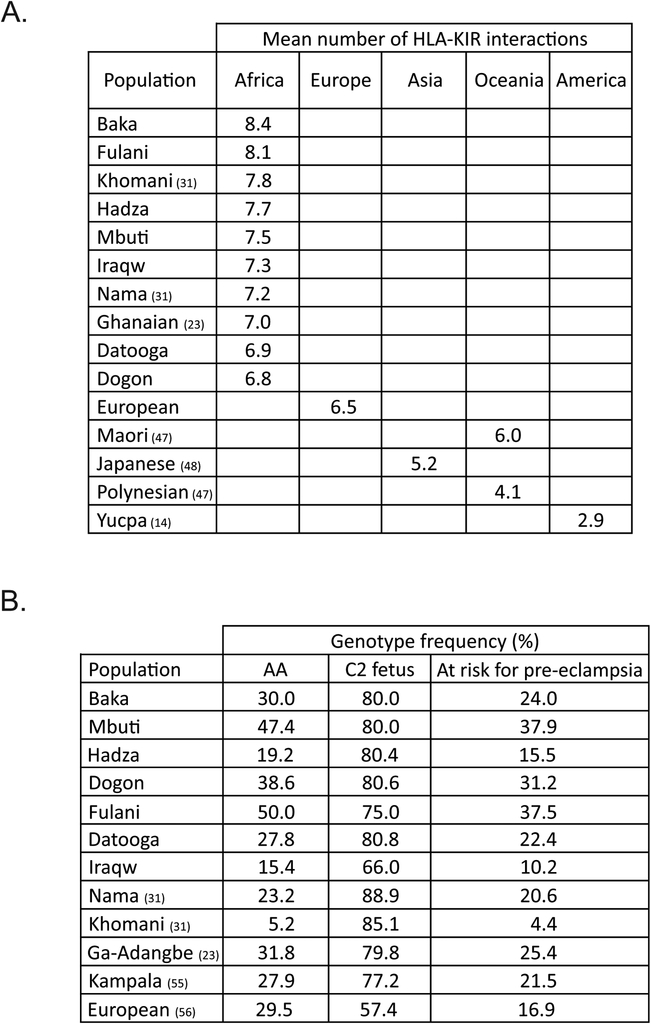
HLA class I and KIR sequences were determined for Dogon, Fulani, and Baka populations of western Africa, Mbuti of central Africa, and Datooga, Iraqw, and Hadza of eastern Africa. Study of 162 individuals identified 134 HLA class I alleles (41 HLA-A, 60 HLA-B, and 33 HLA-C). Common to all populations are three HLA-C alleles (C1+C*07:01, C1+C*07:02, and C2+C*06:02) but no HLA-A or -B Unexpectedly, no novel HLA class I was identified in these previously unstudied and anthropologically distinctive populations. In contrast, of 227 KIR detected, 22 are present in all seven populations and 28 are novel. A high diversity of HLA A-C-B haplotypes was observed. In six populations, most haplotypes are represented just once. But in the Hadza, a majority of haplotypes occur more than once, with 2 having high frequencies and 10 having intermediate frequencies. The centromeric (cen) part of the KIR locus exhibits an even balance between cenA and cenB in all seven populations. The telomeric (tel) part has an even balance of telA to telB in East Africa, but this changes across the continent to where telB is vestigial in West Africa. All four KIR ligands (A3/11, Bw4, C1, and C2) are present in six of the populations. HLA haplotypes of the Iraqw and Hadza encode two KIR ligands, whereas the other populations have an even balance between haplotypes encoding one and two KIR ligands. Individuals in these African populations have a mean of 6.8-8.4 different interactions between KIR and HLA class I, compared with 2.9-6.5 for non-Africans.
Copyright © 2019 by The American Association of Immunologists, Inc.
Figures




References
-
- King A, Boocock C, Sharkey AM, Gardner L, Beretta A, Siccardi AG, and Loke YW. 1996. Evidence for the expression of HLAA-C class I mRNA and protein by human first trimester trophoblast. J. Immunol 156: 2068–2076. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
