

Analysis of Coronavirus Temperature-Sensitive Mutants Reveals an Interplay between the Macrodomain and Papain-Like Protease Impacting Replication and Pathogenesis
- PMID: 30918076
- PMCID: PMC6613754
- DOI: 10.1128/JVI.02140-18
Analysis of Coronavirus Temperature-Sensitive Mutants Reveals an Interplay between the Macrodomain and Papain-Like Protease Impacting Replication and Pathogenesis
Abstract
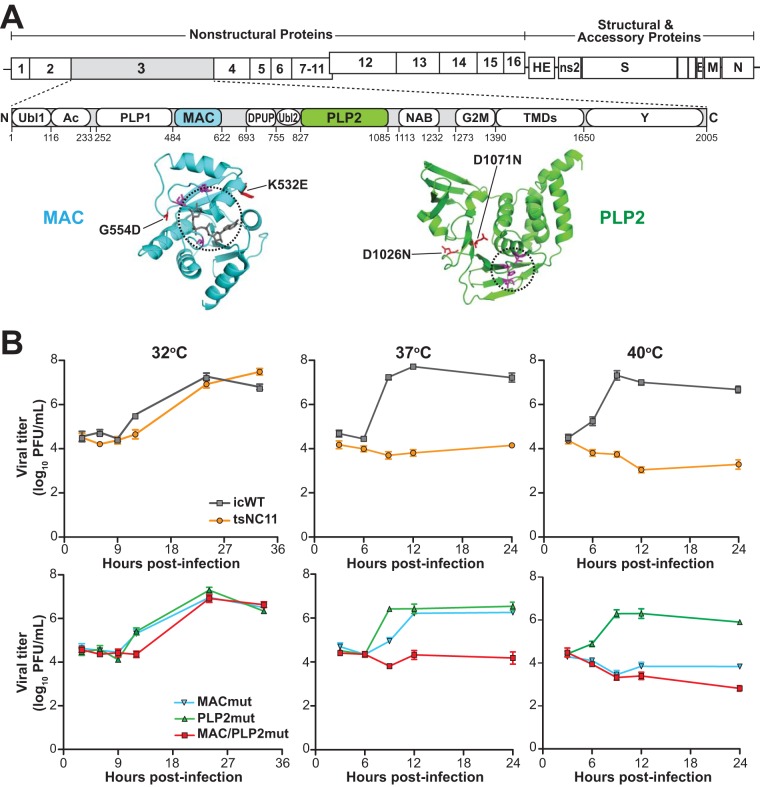
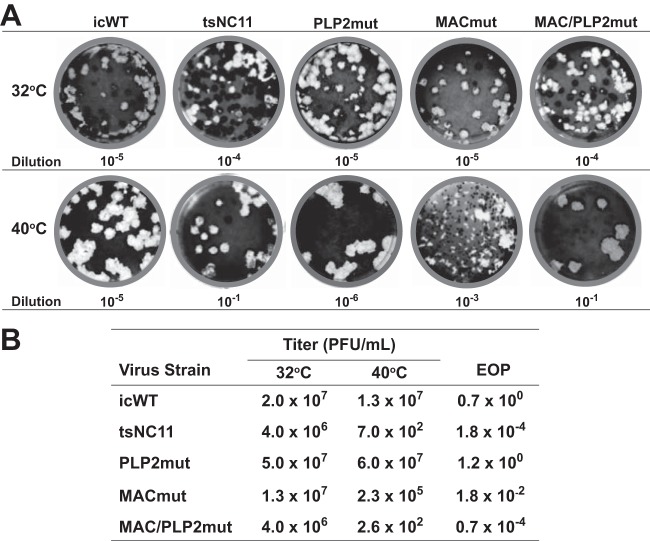
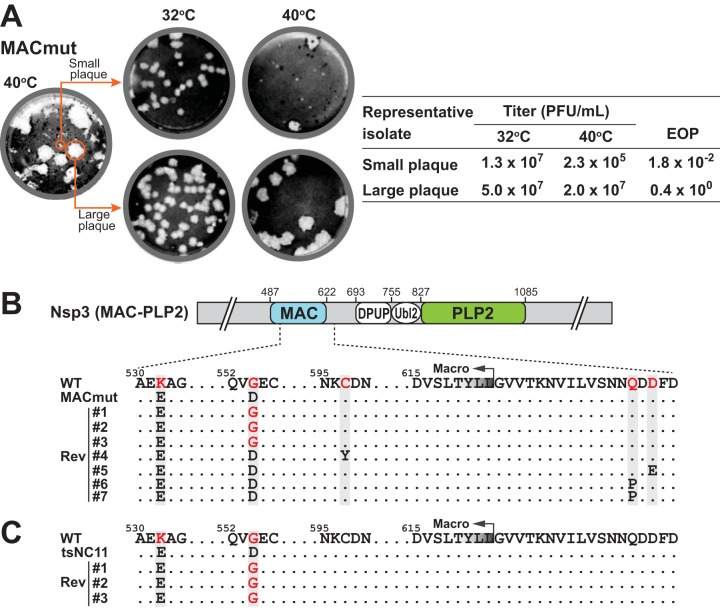
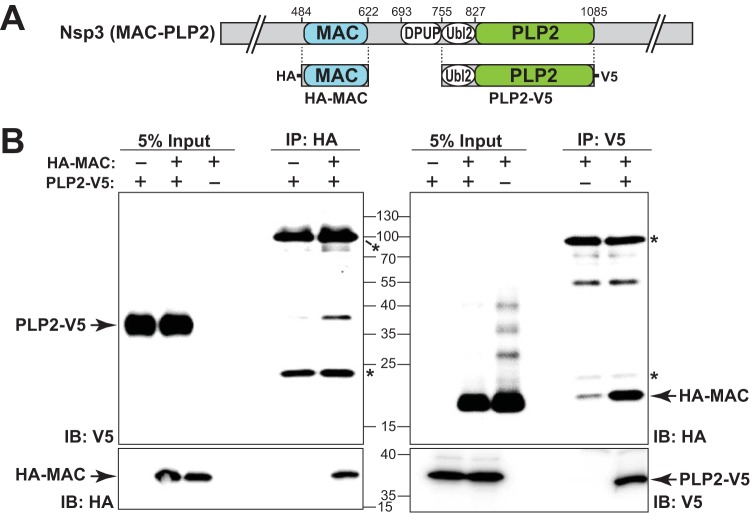
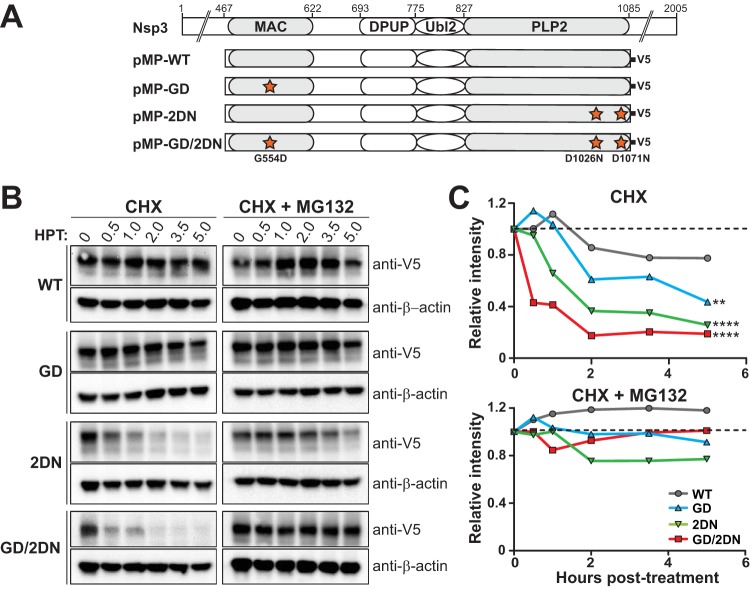
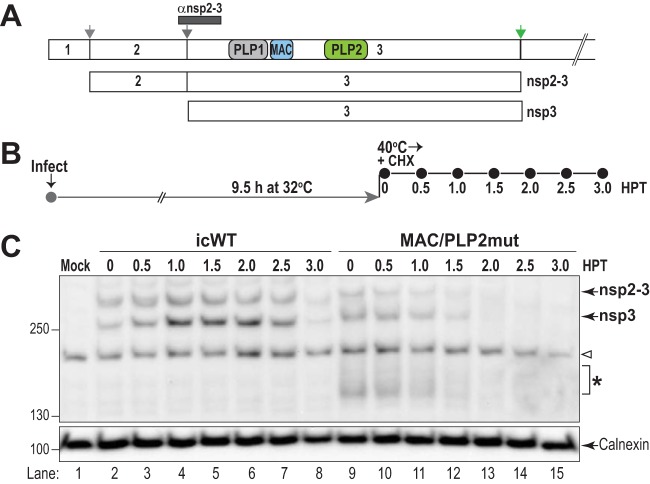
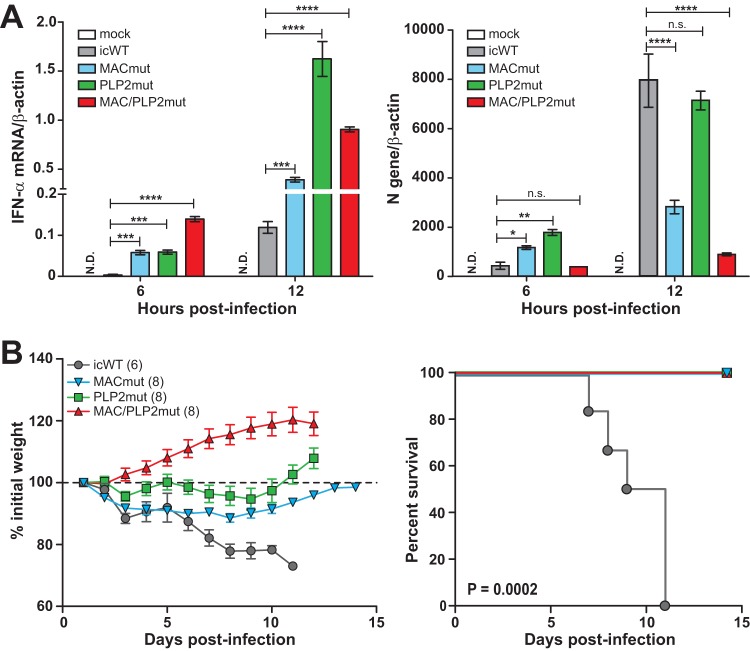
Analysis of temperature-sensitive (ts) mutant viruses is a classic method allowing researchers to identify genetic loci involved in viral replication and pathogenesis. Here, we report genetic analysis of a ts strain of mouse hepatitis virus (MHV), tsNC11, focusing on the role of mutations in the macrodomain (MAC) and the papain-like protease 2 (PLP2) domain of nonstructural protein 3 (nsp3), a component of the viral replication complex. Using MHV reverse genetics, we generated a series of mutant viruses to define the contributions of macrodomain- and PLP2-specific mutations to the ts phenotype. Viral replication kinetics and efficiency-of-plating analysis performed at permissive and nonpermissive temperatures revealed that changes in the macrodomain alone were both necessary and sufficient for the ts phenotype. Interestingly, mutations in the PLP2 domain were not responsible for the temperature sensitivity but did reduce the frequency of reversion of macrodomain mutants. Coimmunoprecipitation studies are consistent with an interaction between the macrodomain and PLP2. Expression studies of the macrodomain-PLP2 portion of nsp3 indicate that the ts mutations enhance proteasome-mediated degradation of the protein. Furthermore, we found that during virus infection, the replicase proteins containing the MAC and PLP2 mutations were more rapidly degraded at the nonpermissive temperature than were the wild-type proteins. Importantly, we show that the macrodomain and PLP2 mutant viruses trigger production of type I interferon in vitro and are attenuated in mice, further highlighting the importance of the macrodomain-PLP2 interplay in viral pathogenesis.IMPORTANCE Coronaviruses (CoVs) are emerging human and veterinary pathogens with pandemic potential. Despite the established and predicted threat these viruses pose to human health, there are currently no approved countermeasures to control infections with these viruses in humans. Viral macrodomains, enzymes that remove posttranslational ADP-ribosylation of proteins, and viral multifunctional papain-like proteases, enzymes that cleave polyproteins and remove polyubiquitin chains via deubiquitinating activity, are two important virulence factors. Here, we reveal an unanticipated interplay between the macrodomain and the PLP2 domain that is important for replication and antagonizing the host innate immune response. Targeting the interaction of these enzymes may provide new therapeutic opportunities to treat CoV disease.
Keywords: coronavirus; innate immunity; interferon; macrodomain; papain-like protease; temperature sensitive; viral replication.
Copyright © 2019 American Society for Microbiology.
Figures
Similar articles
-
Murine coronavirus ubiquitin-like domain is important for papain-like protease stability and viral pathogenesis.J Virol. 2015 May;89(9):4907-17. doi: 10.1128/JVI.00338-15. Epub 2015 Feb 18. J Virol. 2015. PMID: 25694594 Free PMC article.
-
The nsp3 macrodomain promotes virulence in mice with coronavirus-induced encephalitis.J Virol. 2015 Feb;89(3):1523-36. doi: 10.1128/JVI.02596-14. Epub 2014 Nov 26. J Virol. 2015. PMID: 25428866 Free PMC article.
-
Structure-Guided Mutagenesis Alters Deubiquitinating Activity and Attenuates Pathogenesis of a Murine Coronavirus.J Virol. 2020 May 18;94(11):e01734-19. doi: 10.1128/JVI.01734-19. Print 2020 May 18. J Virol. 2020. PMID: 32188728 Free PMC article.
-
Nidovirus papain-like proteases: multifunctional enzymes with protease, deubiquitinating and deISGylating activities.Virus Res. 2014 Dec 19;194:184-90. doi: 10.1016/j.virusres.2014.01.025. Epub 2014 Feb 7. Virus Res. 2014. PMID: 24512893 Free PMC article. Review.
-
Nsp3 of coronaviruses: Structures and functions of a large multi-domain protein.Antiviral Res. 2018 Jan;149:58-74. doi: 10.1016/j.antiviral.2017.11.001. Epub 2017 Nov 8. Antiviral Res. 2018. PMID: 29128390 Free PMC article. Review.
Cited by
-
Replicase 1a gene plays a critical role in pathogenesis of avian coronavirus infectious bronchitis virus.Virology. 2020 Nov;550:1-7. doi: 10.1016/j.virol.2020.08.009. Epub 2020 Aug 20. Virology. 2020. PMID: 32853833 Free PMC article.
-
Insight into the mechanisms of coronaviruses evading host innate immunity.Biochim Biophys Acta Mol Basis Dis. 2023 Jun;1869(5):166671. doi: 10.1016/j.bbadis.2023.166671. Epub 2023 Feb 27. Biochim Biophys Acta Mol Basis Dis. 2023. PMID: 36858323 Free PMC article. Review.
-
Mutation of a highly conserved isoleucine residue in loop 2 of several β-coronavirus macrodomains indicates that enhanced ADP-ribose binding is detrimental for replication.J Virol. 2024 Nov 19;98(11):e0131324. doi: 10.1128/jvi.01313-24. Epub 2024 Oct 10. J Virol. 2024. PMID: 39387584 Free PMC article.
-
The Cold-Adapted, Temperature-Sensitive SARS-CoV-2 Strain TS11 Is Attenuated in Syrian Hamsters and a Candidate Attenuated Vaccine.Viruses. 2022 Dec 29;15(1):95. doi: 10.3390/v15010095. Viruses. 2022. PMID: 36680135 Free PMC article.
-
A persistent high ambient temperature waned the community spread of severe acute respiratory syndrome coronavirus-2 in Pakistan.New Microbes New Infect. 2022 Jan;45:100961. doi: 10.1016/j.nmni.2022.100961. Epub 2022 Jan 24. New Microbes New Infect. 2022. PMID: 35096398 Free PMC article.
References
-
- Zhou P, Fan H, Lan T, Yang X-L, Shi W-F, Zhang W, Zhu Y, Zhang Y-W, Xie Q-M, Mani S, Zheng X-S, Li B, Li J-MJ, Guo H, Pei G-Q, An X-P, Chen JJ-W, Zhou L, Mai K-J, Wu Z-X, Li D, Anderson DE, Zhang L-B, Li S-Y, Mi Z-Q, He T-T, Cong F, Guo P-J, Huang R, Luo Y, Liu X-L, Chen JJ-W, Huang Y, Sun Q, Zhang X-L-L, Wang Y-Y, Xing S-Z, Chen Y-S, Sun Y, Li J-MJ, Daszak P, Wang L-F, Shi Z-L, Tong Y-G, Ma J-Y. 2018. Fatal swine acute diarrhoea syndrome caused by an HKU2-related coronavirus of bat origin. Nature 556:255–258. doi:10.1038/s41586-018-0010-9. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
