

Histone deacetylase inhibitors restore normal hippocampal synaptic plasticity and seizure threshold in a mouse model of Tuberous Sclerosis Complex
- PMID: 30918308
- PMCID: PMC6437206
- DOI: 10.1038/s41598-019-41744-7
Histone deacetylase inhibitors restore normal hippocampal synaptic plasticity and seizure threshold in a mouse model of Tuberous Sclerosis Complex
Abstract
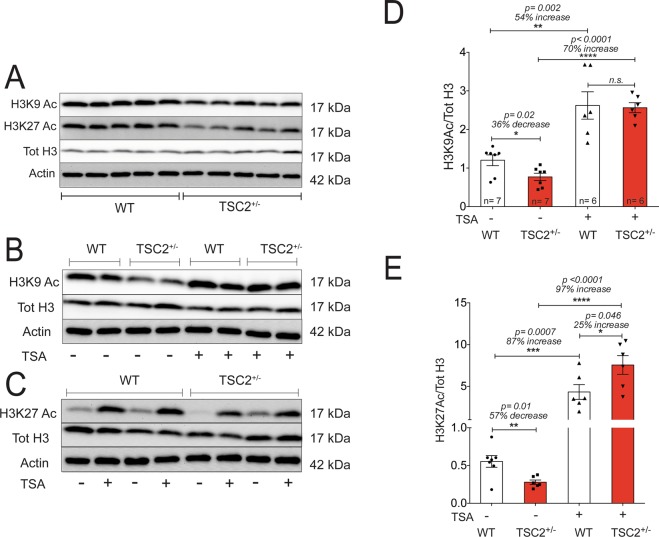
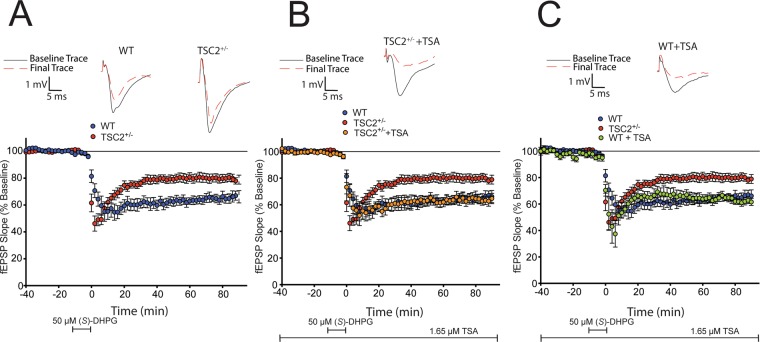
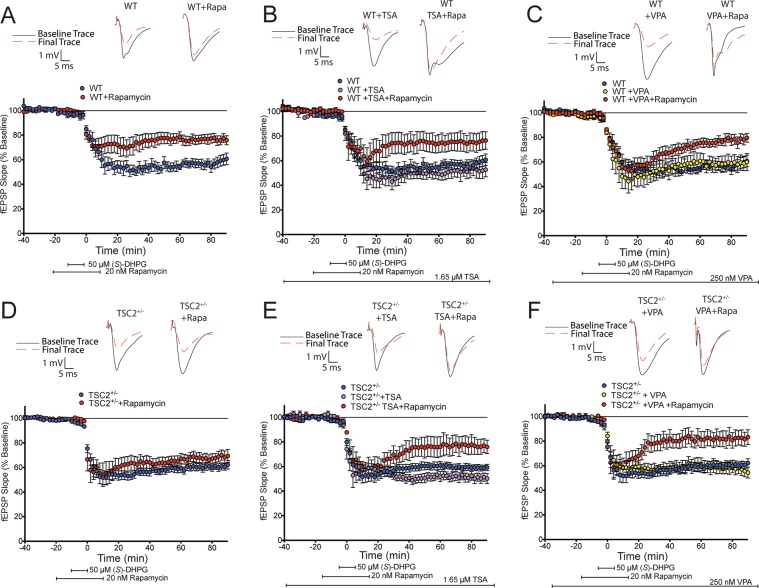
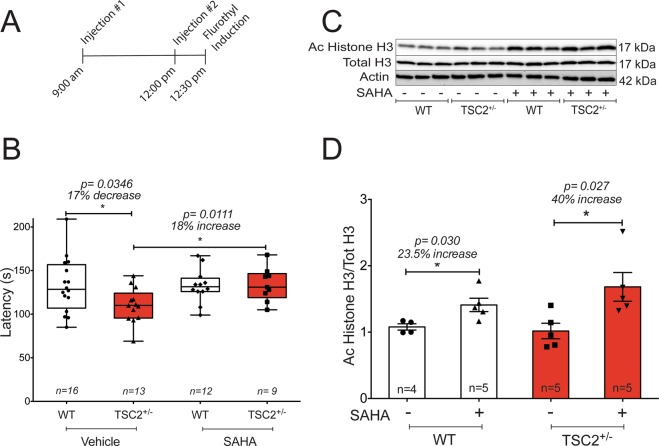
Abnormal synaptic plasticity has been implicated in several neurological disorders including epilepsy, dementia and Autism Spectrum Disorder (ASD). Tuberous Sclerosis Complex (TSC) is an autosomal dominant genetic disorder that manifests with seizures, autism, and cognitive deficits. The abnormal intracellular signaling underlying TSC has been the focus of many studies. However, nothing is known about the role of histone modifications in contributing to the neurological manifestations in TSC. Dynamic regulation of chromatin structure via post translational modification of histone tails has been implicated in learning, memory and synaptic plasticity. Histone acetylation and associated gene activation plays a key role in plasticity and so we asked whether histone acetylation might be dysregulated in TSC. In this study, we report a general reduction in hippocampal histone H3 acetylation levels in a mouse model of TSC2. Pharmacological inhibition of Histone Deacetylase (HDAC) activity restores histone H3 acetylation levels and ameliorates the aberrant plasticity in TSC2+/- mice. We describe a novel seizure phenotype in TSC2+/- mice that is also normalized with HDAC inhibitors (HDACis). The results from this study suggest an unanticipated role for chromatin modification in TSC and may inform novel therapeutic strategies for TSC patients.
Conflict of interest statement
The authors declare no competing interests.
Figures

Comment in
-
Epigenetic Therapeutic Intervention for a Rare Epilepsy Disorder.Epilepsy Curr. 2020 Mar;20(2):111-112. doi: 10.1177/1535759720904162. Epub 2020 Feb 17. Epilepsy Curr. 2020. PMID: 32064921 Free PMC article.
References
-
- Martin, S., Grimwood, P. & Morris, R. Synaptic Plasticity and Memory: An Evaluation of the Hypothesis. Annual review of neuroscience 649–711, 10.1146/annurev.neuro.23.1.649 (2000). - PubMed
-
- Hebb, D. O. The organization of Behavior. John Wiley & Sons, Inc. (John Wiley & Sons, Inc., 1949).
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
