

Agent-based modeling of morphogenetic systems: Advantages and challenges
- PMID: 30921323
- PMCID: PMC6438454
- DOI: 10.1371/journal.pcbi.1006577
Agent-based modeling of morphogenetic systems: Advantages and challenges
Abstract
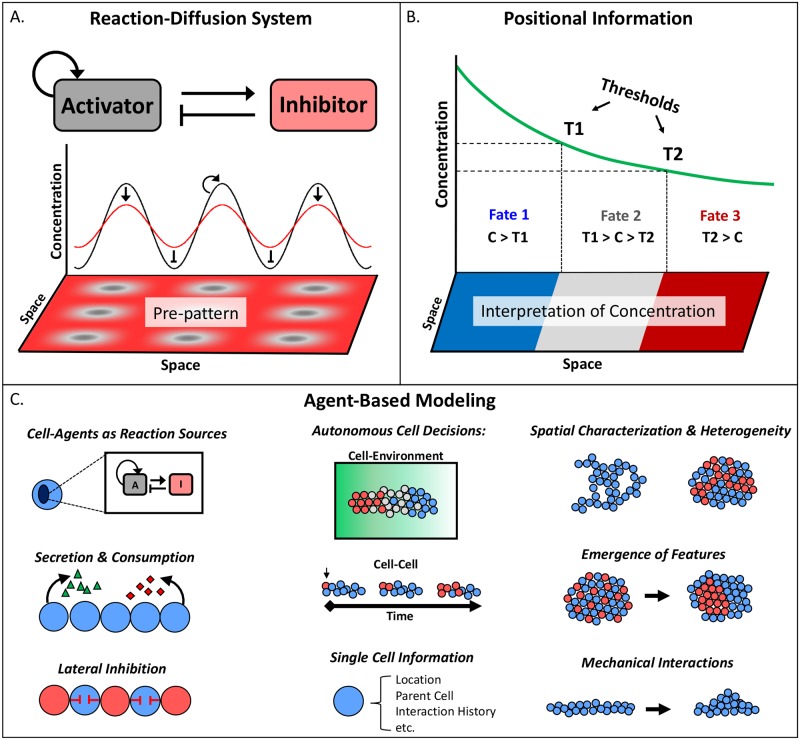
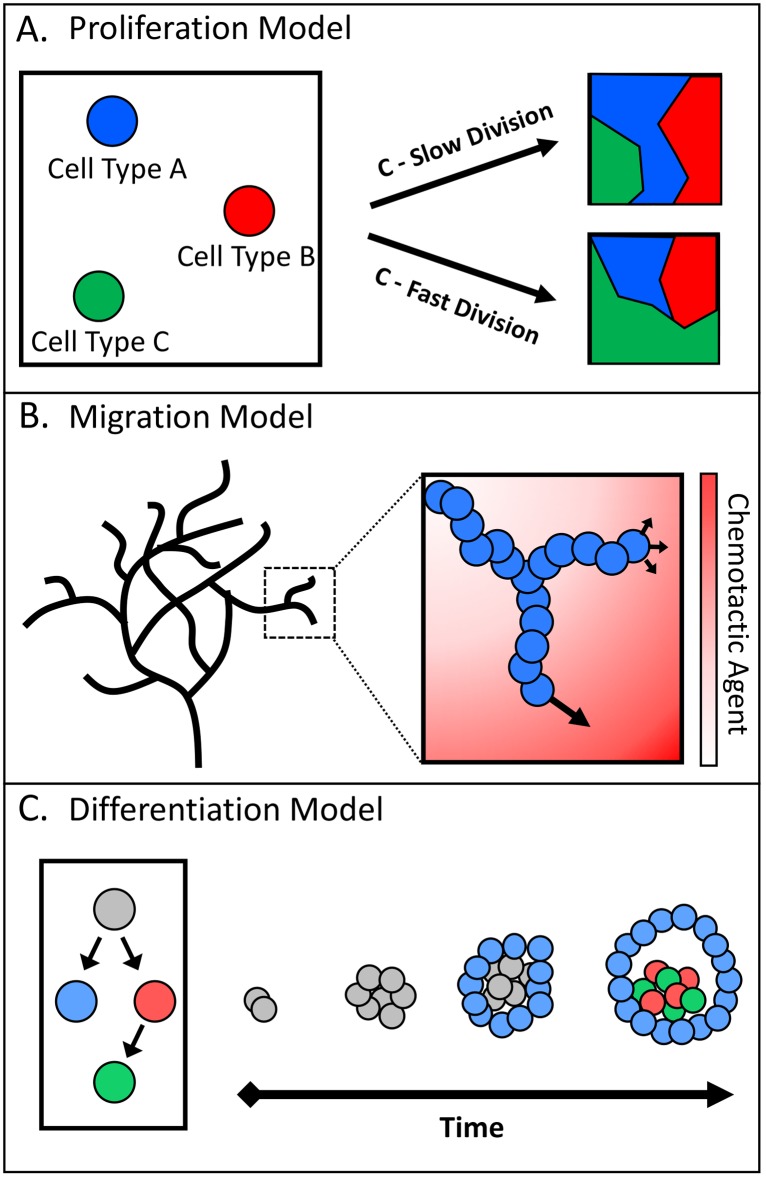
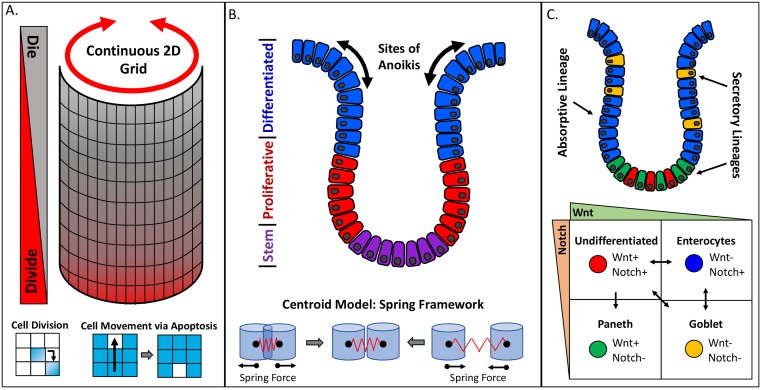
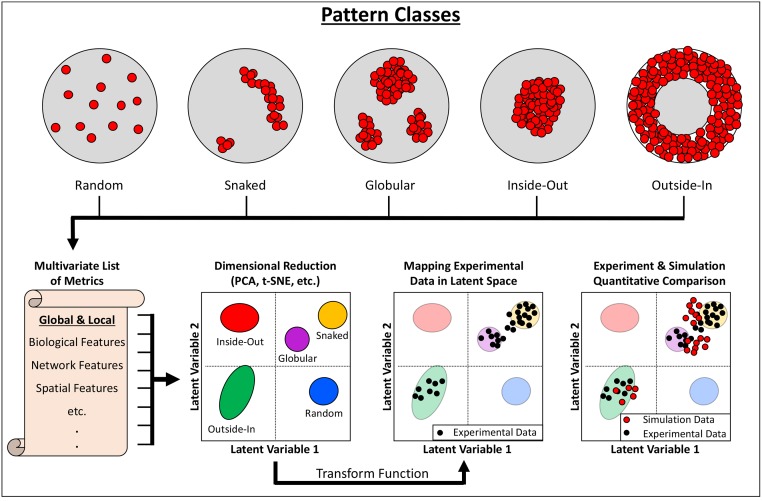
The complexity of morphogenesis poses a fundamental challenge to understanding the mechanisms governing the formation of biological patterns and structures. Over the past century, numerous processes have been identified as critically contributing to morphogenetic events, but the interplay between the various components and aspects of pattern formation have been much harder to grasp. The combination of traditional biology with mathematical and computational methods has had a profound effect on our current understanding of morphogenesis and led to significant insights and advancements in the field. In particular, the theoretical concepts of reaction-diffusion systems and positional information, proposed by Alan Turing and Lewis Wolpert, respectively, dramatically influenced our general view of morphogenesis, although typically in isolation from one another. In recent years, agent-based modeling has been emerging as a consolidation and implementation of the two theories within a single framework. Agent-based models (ABMs) are unique in their ability to integrate combinations of heterogeneous processes and investigate their respective dynamics, especially in the context of spatial phenomena. In this review, we highlight the benefits and technical challenges associated with ABMs as tools for examining morphogenetic events. These models display unparalleled flexibility for studying various morphogenetic phenomena at multiple levels and have the important advantage of informing future experimental work, including the targeted engineering of tissues and organs.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

Similar articles
-
On the evolution of morphogenetic models: mechano-chemical interactions and an integrated view of cell differentiation, growth, pattern formation and morphogenesis.Biol Rev Camb Philos Soc. 2012 Nov;87(4):786-803. doi: 10.1111/j.1469-185X.2012.00221.x. Epub 2012 Mar 19. Biol Rev Camb Philos Soc. 2012. PMID: 22429266 Review.
-
Insights from chemical systems into Turing-type morphogenesis.Philos Trans A Math Phys Eng Sci. 2021 Dec 27;379(2213):20200269. doi: 10.1098/rsta.2020.0269. Epub 2021 Nov 8. Philos Trans A Math Phys Eng Sci. 2021. PMID: 34743602 Review.
-
Modern perspectives on near-equilibrium analysis of Turing systems.Philos Trans A Math Phys Eng Sci. 2021 Dec 27;379(2213):20200268. doi: 10.1098/rsta.2020.0268. Epub 2021 Nov 8. Philos Trans A Math Phys Eng Sci. 2021. PMID: 34743603 Free PMC article. Review.
-
A general formalism for tissue morphogenesis based on cellular dynamics and control system interactions.Acta Biotheor. 2008 Jun;56(1-2):51-74. doi: 10.1007/s10441-008-9030-4. Epub 2008 Feb 15. Acta Biotheor. 2008. PMID: 18274863
-
Mechanocellular models of epithelial morphogenesis.Philos Trans R Soc Lond B Biol Sci. 2017 May 19;372(1720):20150519. doi: 10.1098/rstb.2015.0519. Philos Trans R Soc Lond B Biol Sci. 2017. PMID: 28348253 Free PMC article. Review.
Cited by
-
Calibrating agent-based models to tumor images using representation learning.PLoS Comput Biol. 2023 Apr 21;19(4):e1011070. doi: 10.1371/journal.pcbi.1011070. eCollection 2023 Apr. PLoS Comput Biol. 2023. PMID: 37083821 Free PMC article.
-
Toward Engineering Biosystems With Emergent Collective Functions.Front Bioeng Biotechnol. 2020 Jun 26;8:705. doi: 10.3389/fbioe.2020.00705. eCollection 2020. Front Bioeng Biotechnol. 2020. PMID: 32671054 Free PMC article.
-
Computational Biology and in silico Toxicodynamics.Curr Opin Toxicol. 2020 Dec 1;23-24(Oct-Dec 2020):119-126. doi: 10.1016/j.cotox.2020.11.001. Curr Opin Toxicol. 2020. PMID: 36561131 Free PMC article.
-
Engineering multicellular living systems-a Keystone Symposia report.Ann N Y Acad Sci. 2022 Dec;1518(1):183-195. doi: 10.1111/nyas.14896. Epub 2022 Sep 30. Ann N Y Acad Sci. 2022. PMID: 36177947 Free PMC article.
-
Morphogenetic metasurfaces: unlocking the potential of turing patterns.Nat Commun. 2023 Oct 6;14(1):6249. doi: 10.1038/s41467-023-41775-9. Nat Commun. 2023. PMID: 37803018 Free PMC article.
References
-
- Thompson DW. On Growth and Form. Cambridge, UK: Cambridge University Press; 1942.
-
- Turing AM. The Chemical Basis of Morphogenesis. Phil Trans R Soc Lond. 1952;237(641):37–72.
-
- Segel LA, Jackson JL. Dissipative Structure: An Explanation and an Ecological Example. J Theor Biol. 1972;37:545–59. - PubMed
-
- Meinhardt H, Gierer A. Applications of a Theory of Biological Pattern Formation Based on Lateral Inhibition. J Cell Science. 1974;15:321–46. - PubMed
-
- Wolpert L. Chapter 6 Positional Information and Pattern Formation. Current Topics in Developmental Biology. 1971;6:183–224. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous
