

Rice aquaporin PIP1;3 and harpin Hpa1 of bacterial blight pathogen cooperate in a type III effector translocation
- PMID: 30921464
- PMCID: PMC6598099
- DOI: 10.1093/jxb/erz130
Rice aquaporin PIP1;3 and harpin Hpa1 of bacterial blight pathogen cooperate in a type III effector translocation
Abstract
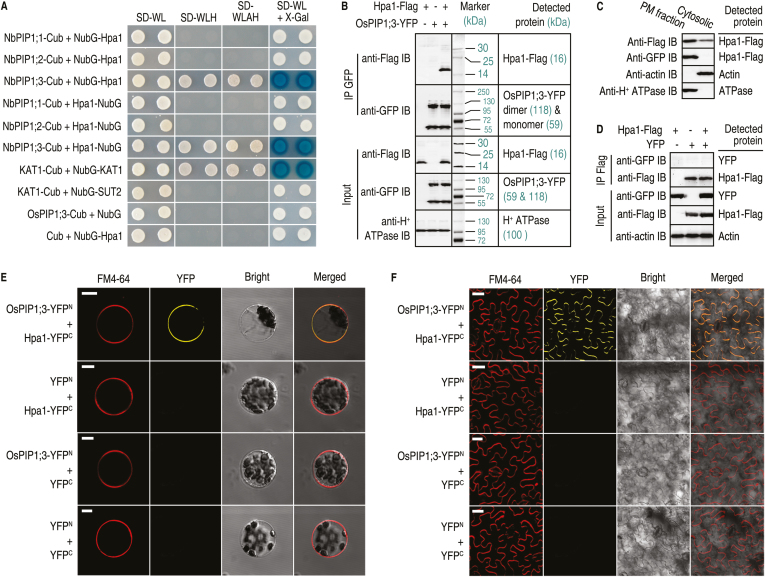
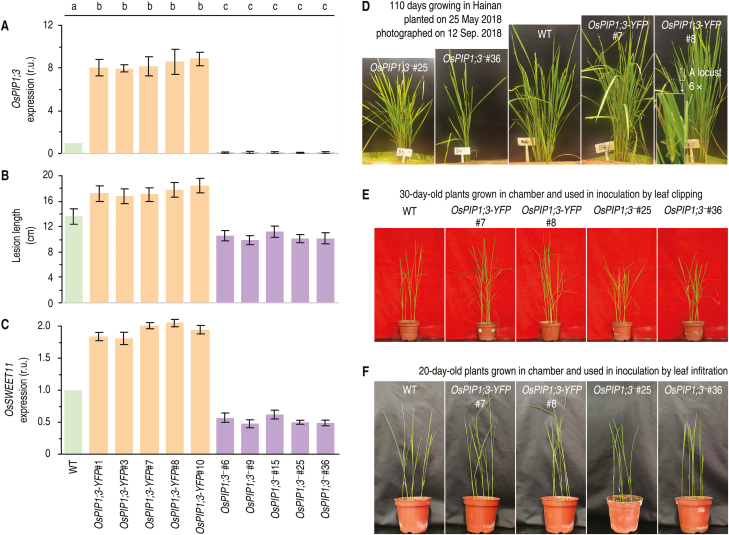
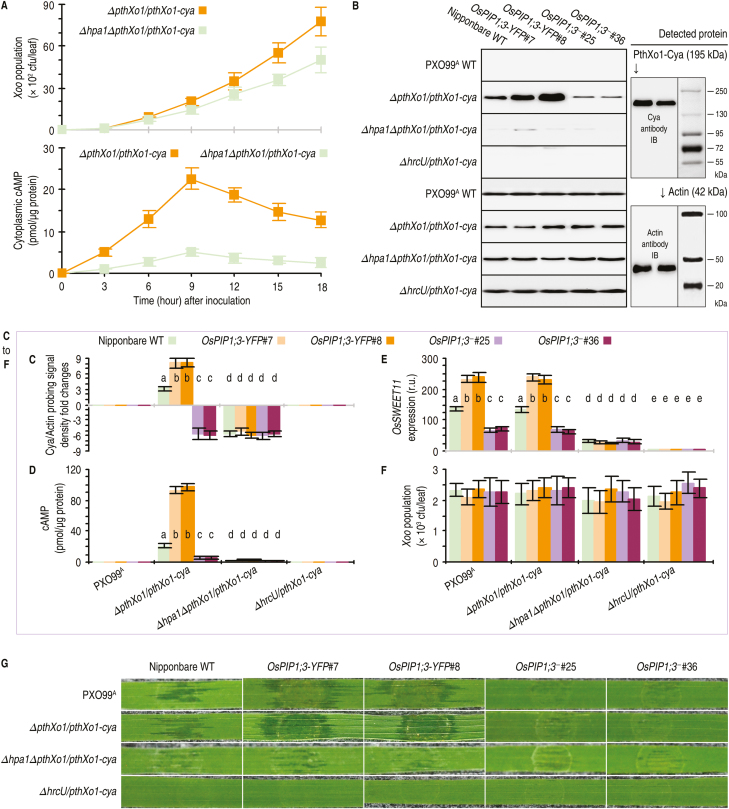
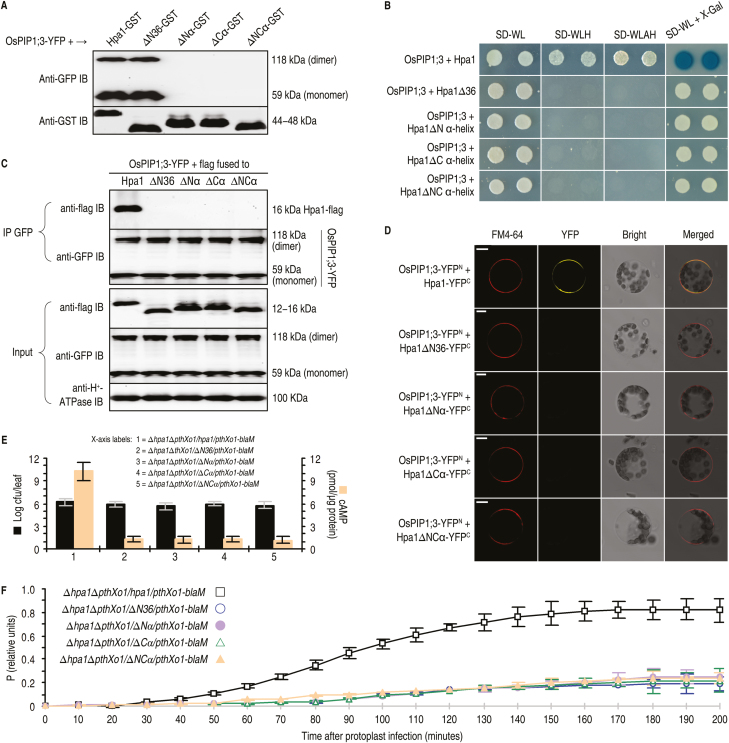
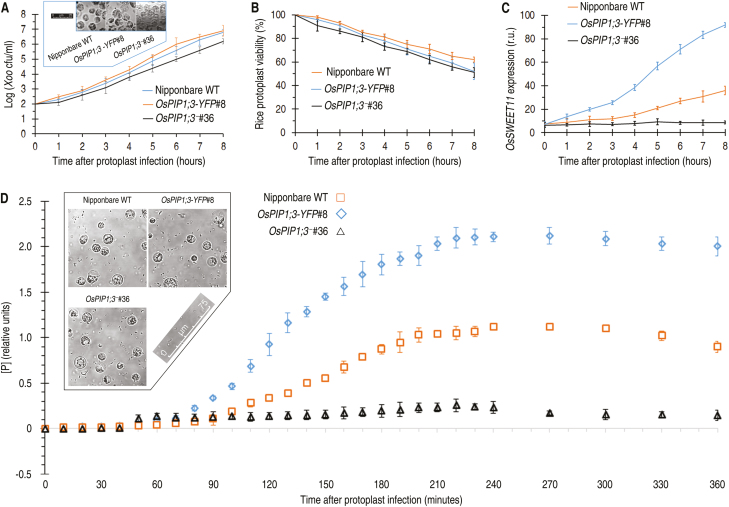
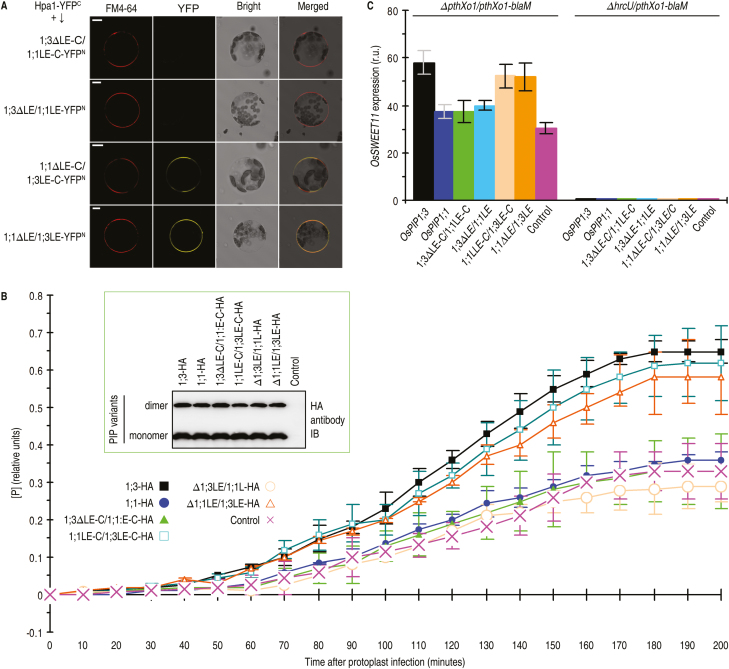
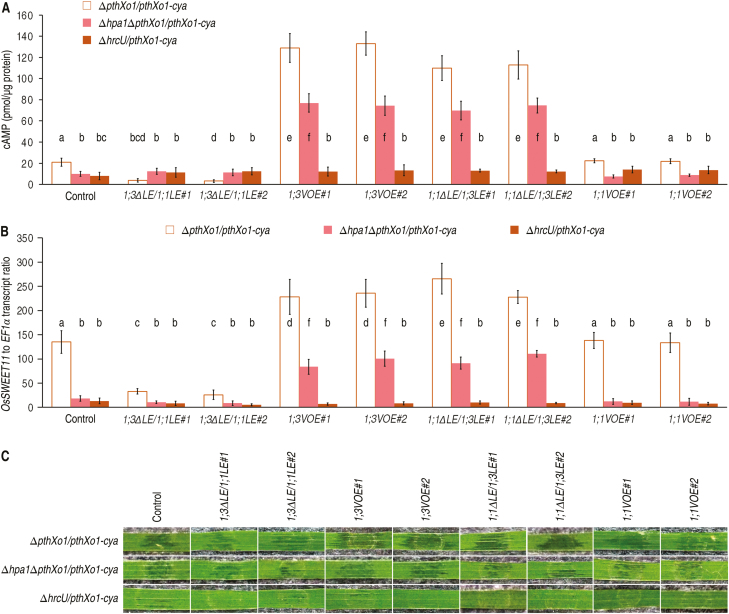
Varieties of Gram-negative bacterial pathogens infect their eukaryotic hosts by deploying the type III translocon to deliver effector proteins into the cytosol of eukaryotic cells in which effectors execute their pathological functions. The translocon is hypothetically assembled by bacterial translocators in association with the assumed receptors situated on eukaryotic plasma membranes. This hypothesis is partially verified in the present study with genetic, biochemical, and pathological evidence for the role of a rice aquaporin, plasma membrane intrinsic protein PIP1;3, in the cytosolic import of the transcription activator-like effector PthXo1 from the bacterial blight pathogen. PIP1;3 interacts with the bacterial translocator Hpa1 at rice plasma membranes to control PthXo1 translocation from cells of a well-characterized strain of the bacterial blight pathogen into the cytosol of cells of a susceptible rice variety. An extracellular loop sequence of PIP1;3 and the α-helix motif of Hpa1 determine both the molecular interaction and its consequences with respect to the effector translocation and the bacterial virulence on the susceptible rice variety. Overall, these results provide multiple experimental avenues to support the hypothesis that interactions between bacterial translocators and their interactors at the target membrane are essential for bacterial effector translocation.
Keywords: Aquaporin; PIP1; bacterial pathogen; rice; translocation; type III effector.
© The Author(s) 2019. Published by Oxford University Press on behalf of the Society for Experimental Biology.
Figures


References
-
- Alfano JR, Collmer A. 2004. Type III secretion system effector proteins: double agents in bacterial disease and plant defense. Annual Review of Phytopathology 42, 385–414. - PubMed
-
- Bogdanove AJ, Voytas DF. 2011. TAL effectors: customizable proteins for DNA targeting. Science 333, 1843–1846. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
