

Interaction with hyaluronan matrix and miRNA cargo as contributors for in vitro potential of mesenchymal stem cell-derived extracellular vesicles in a model of human osteoarthritic synoviocytes
- PMID: 30922413
- PMCID: PMC6440078
- DOI: 10.1186/s13287-019-1215-z
Interaction with hyaluronan matrix and miRNA cargo as contributors for in vitro potential of mesenchymal stem cell-derived extracellular vesicles in a model of human osteoarthritic synoviocytes
Abstract
Background: Osteoarthritis (OA) is the most prevalent joint disease, and to date, no options for effective tissue repair and restoration are available. With the aim of developing new therapies, the impact of mesenchymal stem cells (MSCs) has been explored, and the efficacy of MSCs started to be deciphered. A strong paracrine capacity relying on both secreted and vesicle-embedded (EVs) protein or nucleic acid-based factors has been proposed as the principal mechanism that contributes to tissue repair. This work investigated the mechanism of internalization of extracellular vesicles (EVs) released by adipose-derived MSCs (ASCs) and the role of shuttled miRNAs in the restoration of homeostasis in an in vitro model of human fibroblast-like synoviocytes (FLSs) from OA patients.
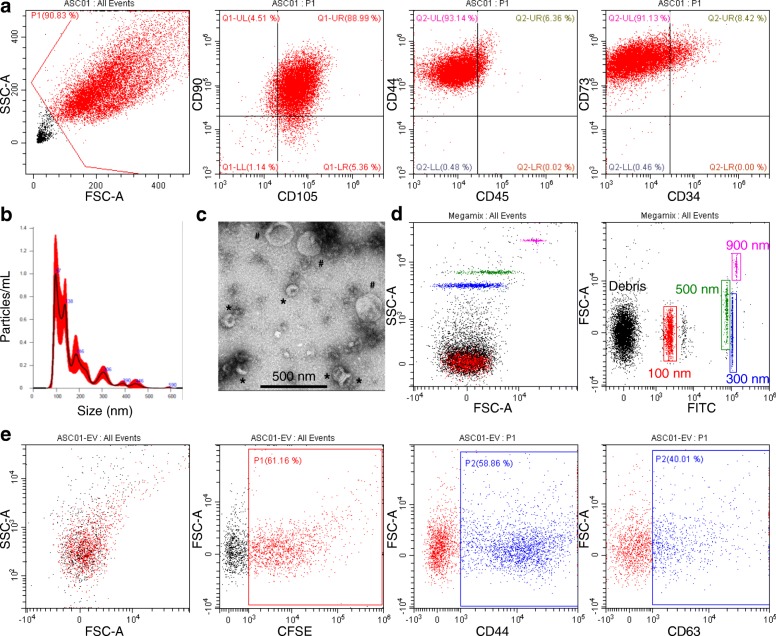
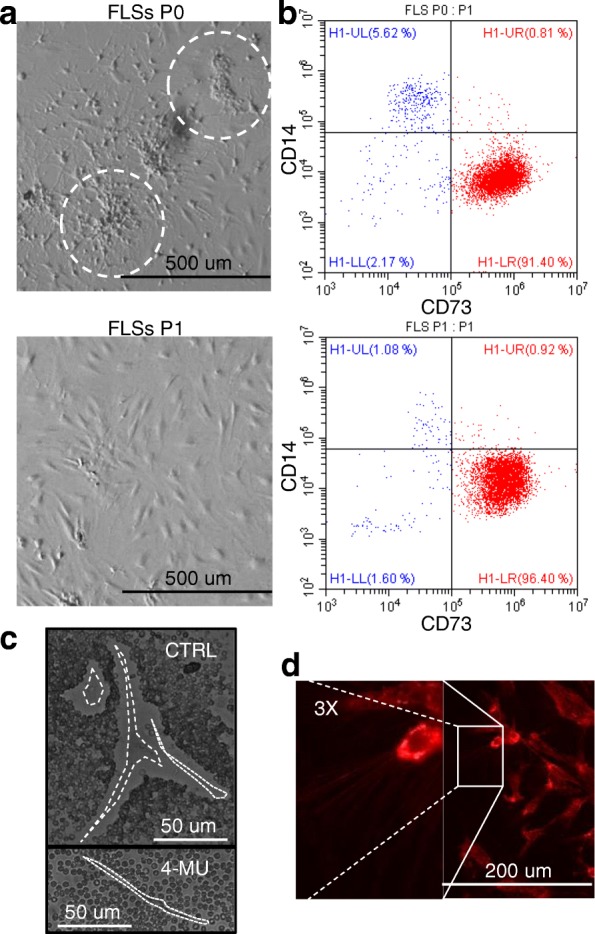
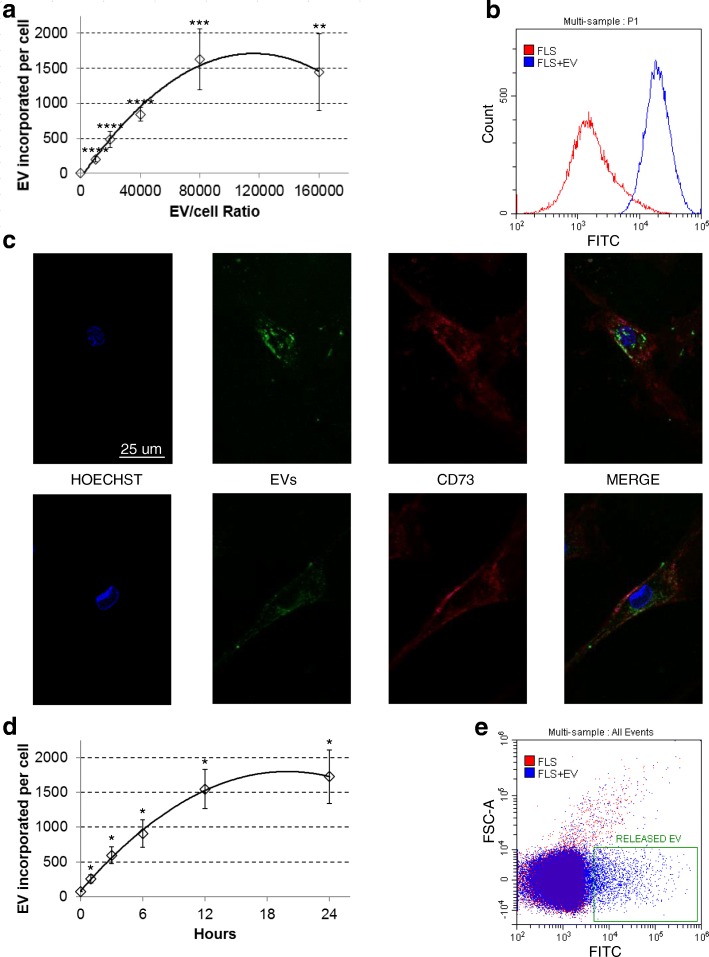
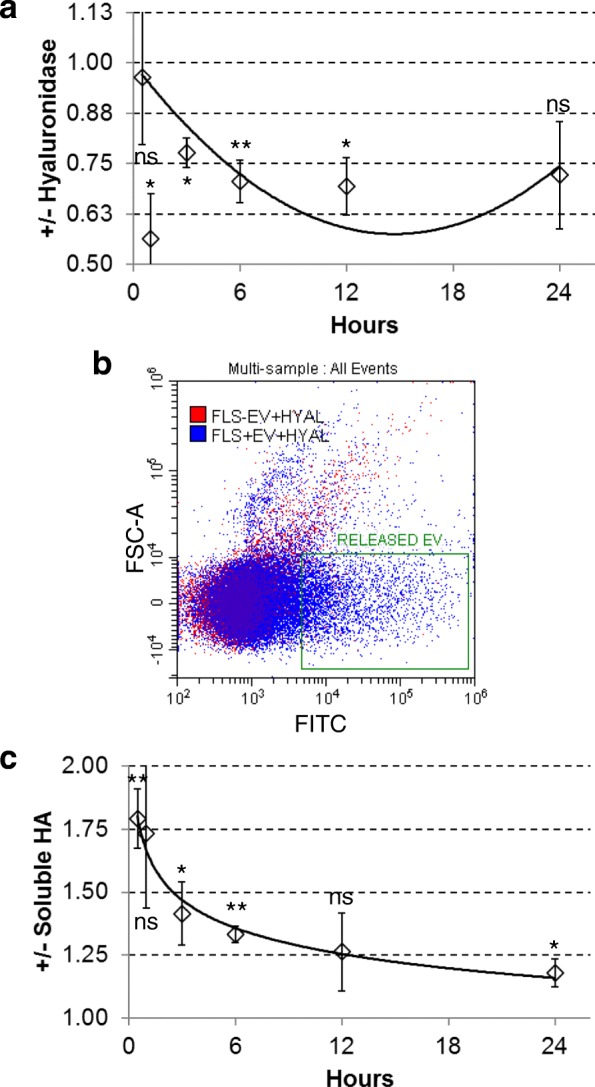
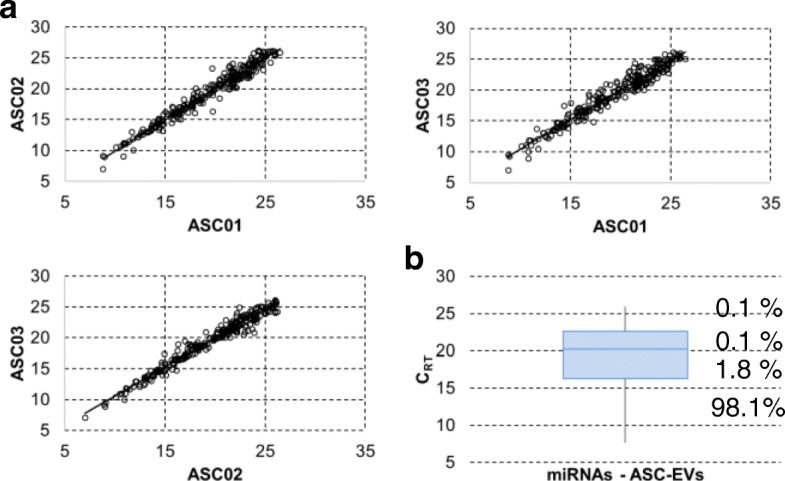
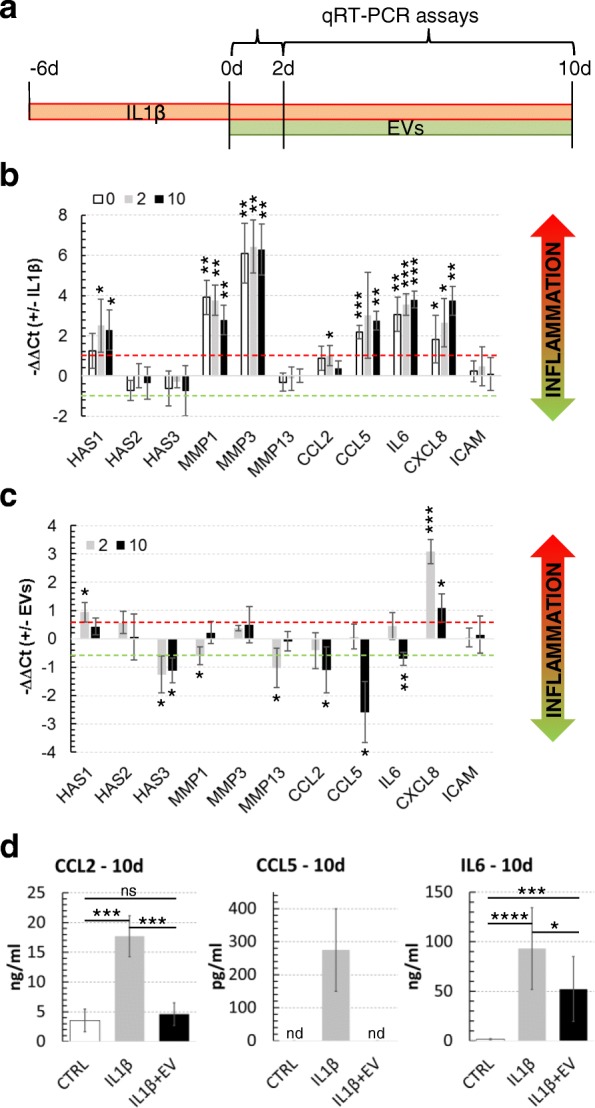
Methods: ASC-EVs were isolated by differential centrifugation and validated by flow cytometry and nanoparticle tracking analysis. ASC-EVs with increased hyaluronan (HA) receptor CD44 levels were obtained culturing ASCs on HA-coated plastic surfaces. OA FLSs with intact or digested HA matrix were co-cultured with fluorescent ASC-EVs, and incorporation scored by flow cytometry and ELISA. ASC-EV complete miRNome was deciphered by high-throughput screening. In inflamed OA FLSs, genes and pathways potentially regulated by ASC-EV miRNA were predicted by bioinformatics. OA FLSs stimulated with IL-1β at physiological levels (25 pg/mL) were treated with ASC-EVs, and expression of inflammation and OA-related genes was measured by qRT-PCR over a 10-day time frame with modulated candidates verified by ELISA.
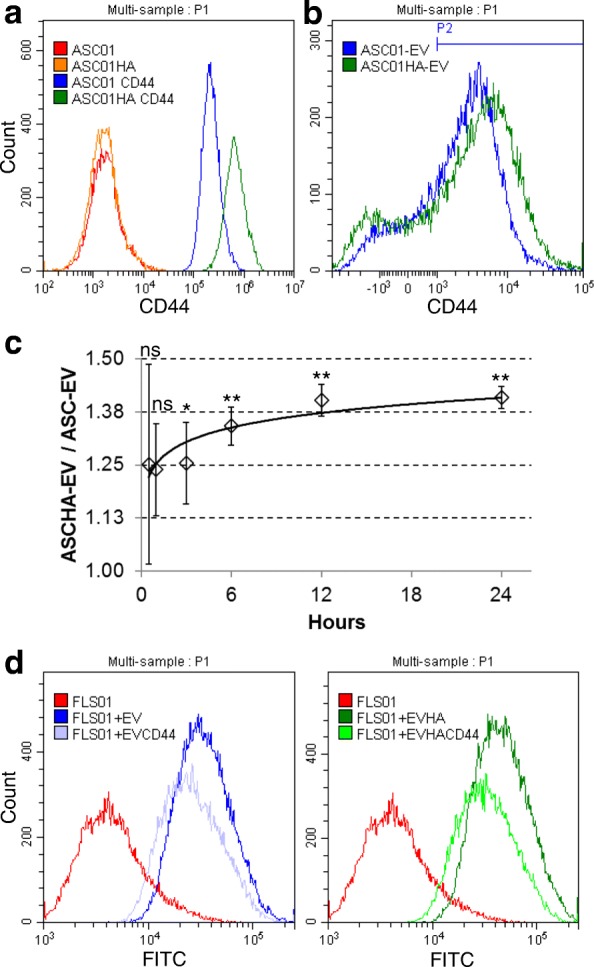
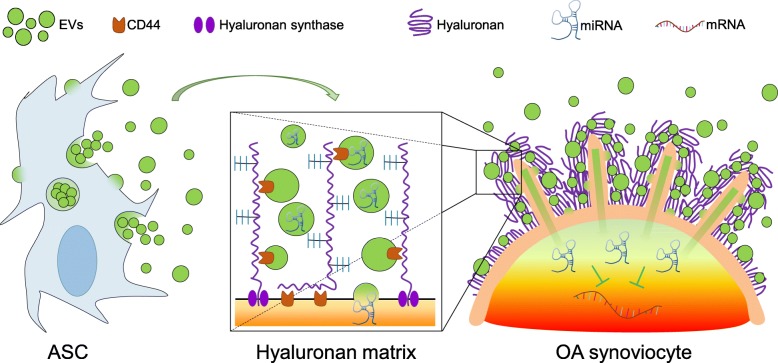
Results: The data showed that HA is involved in ASC-EV internalization in FLSs. Indeed, both removal of HA matrix presence on FLSs and modulation of CD44 levels on EVs affected their recruitment. Bioinformatics analysis of EV-embedded miRNAs showed their ability to potentially regulate the main pathways strictly associated with synovial inflammation in OA. In this frame, ASC-EVs reduced the expression of pro-inflammatory cytokines and chemokines in a chronic model of FLS inflammation.
Conclusions: Given their ability to affect FLS behavior in a model of chronic inflammation through direct interaction with HA matrix and miRNA release, ASC-EVs confirm their role as a novel therapeutic option for osteoarthritic joints.
Keywords: Adipose-derived mesenchymal stem cells; Extracellular vesicles; Hyaluronan coat; Osteoarthritis; Synoviocytes; miRNA.
Conflict of interest statement
Ethics approval and consent to participate
The study was carried out at IRCCS Istituto Ortopedico Galeazzi with Institutional Review Board approval (M-SPER-015 - Ver. 2 - 04.11.2016) and all specimens, both synovial membrane and adipose tissue, were collected with patient informed consent. The experiments comply with the current laws of Italy.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
Similar articles
-
Extracellular Vesicle-Mediated miR-150-3p Delivery in Joint Homeostasis: A Potential Treatment for Osteoarthritis?Cells. 2022 Sep 5;11(17):2766. doi: 10.3390/cells11172766. Cells. 2022. PMID: 36078172 Free PMC article.
-
Adipose-Derived Mesenchymal Stromal Cells Treated with Interleukin 1 Beta Produced Chondro-Protective Vesicles Able to Fast Penetrate in Cartilage.Cells. 2021 May 12;10(5):1180. doi: 10.3390/cells10051180. Cells. 2021. PMID: 34066077 Free PMC article.
-
Inflammatory priming enhances mesenchymal stromal cell secretome potential as a clinical product for regenerative medicine approaches through secreted factors and EV-miRNAs: the example of joint disease.Stem Cell Res Ther. 2020 Apr 28;11(1):165. doi: 10.1186/s13287-020-01677-9. Stem Cell Res Ther. 2020. PMID: 32345351 Free PMC article.
-
Pathogenic or Therapeutic Extracellular Vesicles in Rheumatic Diseases: Role of Mesenchymal Stem Cell-Derived Vesicles.Int J Mol Sci. 2017 Apr 22;18(4):889. doi: 10.3390/ijms18040889. Int J Mol Sci. 2017. PMID: 28441721 Free PMC article. Review.
-
Extracellular vesicles in the pathogenesis of rheumatoid arthritis and osteoarthritis.Arthritis Res Ther. 2016 Dec 1;18(1):286. doi: 10.1186/s13075-016-1178-8. Arthritis Res Ther. 2016. PMID: 27906035 Free PMC article. Review.
Cited by
-
Superior Osteo-Inductive and Osteo-Conductive Properties of Trabecular Titanium vs. PEEK Scaffolds on Human Mesenchymal Stem Cells: A Proof of Concept for the Use of Fusion Cages.Int J Mol Sci. 2021 Feb 27;22(5):2379. doi: 10.3390/ijms22052379. Int J Mol Sci. 2021. PMID: 33673509 Free PMC article.
-
Fibroblast growth factor receptor 1-bound extracellular vesicle as novel therapy for osteoarthritis.Biomedicine (Taipei). 2022 Jun 1;12(2):1-9. doi: 10.37796/2211-8039.1308. eCollection 2022. Biomedicine (Taipei). 2022. PMID: 35836973 Free PMC article. Review.
-
Extracellular vesicles derived from different sources of mesenchymal stem cells: therapeutic effects and translational potential.Cell Biosci. 2020 May 24;10:69. doi: 10.1186/s13578-020-00427-x. eCollection 2020. Cell Biosci. 2020. PMID: 32483483 Free PMC article. Review.
-
Cell-based therapies have disease-modifying effects on osteoarthritis in animal models. A systematic review by the ESSKA Orthobiologic Initiative. Part 1: adipose tissue-derived cell-based injectable therapies.Knee Surg Sports Traumatol Arthrosc. 2023 Feb;31(2):641-655. doi: 10.1007/s00167-022-07063-7. Epub 2022 Sep 14. Knee Surg Sports Traumatol Arthrosc. 2023. PMID: 36104484 Free PMC article.
-
Role of Microparticles in the Pathogenesis of Inflammatory Joint Diseases.Int J Mol Sci. 2019 Nov 1;20(21):5453. doi: 10.3390/ijms20215453. Int J Mol Sci. 2019. PMID: 31683793 Free PMC article. Review.
References
-
- Division UNDoEaSAP. World Population Prospects: The 2015 Revision, Key Findings and Advance Tables 2016. https://esa.un.org/unpd/wpp/. Accessed 19 Dec 2018.
-
- Rannou F, Pelletier JP, Martel-Pelletier J. Efficacy and safety of topical NSAIDs in the management of osteoarthritis: evidence from real-life setting trials and surveys. Semin Arthritis Rheum. 2016. 10.1016/j.semarthrit.2015.11.007. - PubMed
-
- Crisan M, Yap S, Casteilla L, et al. A perivascular origin for mesenchymal stem cells in multiple human organs. Cell Stem Cell. 2008. 10.1016/j.stem.2008.07.003. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
