

Mechanical loading inhibits cartilage inflammatory signalling via an HDAC6 and IFT-dependent mechanism regulating primary cilia elongation
- PMID: 30922983
- PMCID: PMC6593179
- DOI: 10.1016/j.joca.2019.03.003
Mechanical loading inhibits cartilage inflammatory signalling via an HDAC6 and IFT-dependent mechanism regulating primary cilia elongation
Abstract
Objective: Physiological mechanical loading reduces inflammatory signalling in numerous cell types including articular chondrocytes however the mechanism responsible remains unclear. This study investigates the role of chondrocyte primary cilia and associated intraflagellar transport (IFT) in the mechanical regulation of interleukin-1β (IL-1β) signalling.
Design: Isolated chondrocytes and cartilage explants were subjected to cyclic mechanical loading in the presence and absence of the cytokine IL-1β. Nitric oxide (NO) and prostaglandin E2 (PGE2) release were used to monitor IL-1β signalling whilst Sulphated glycosaminoglycan (sGAG) release provided measurement of cartilage degradation. Measurements were made of HDAC6 activity and tubulin polymerisation and acetylation. Effects on primary cilia were monitored by confocal and super resolution microscopy. Involvement of IFT was analysed using ORPK cells with hypomorphic mutation of IFT88.
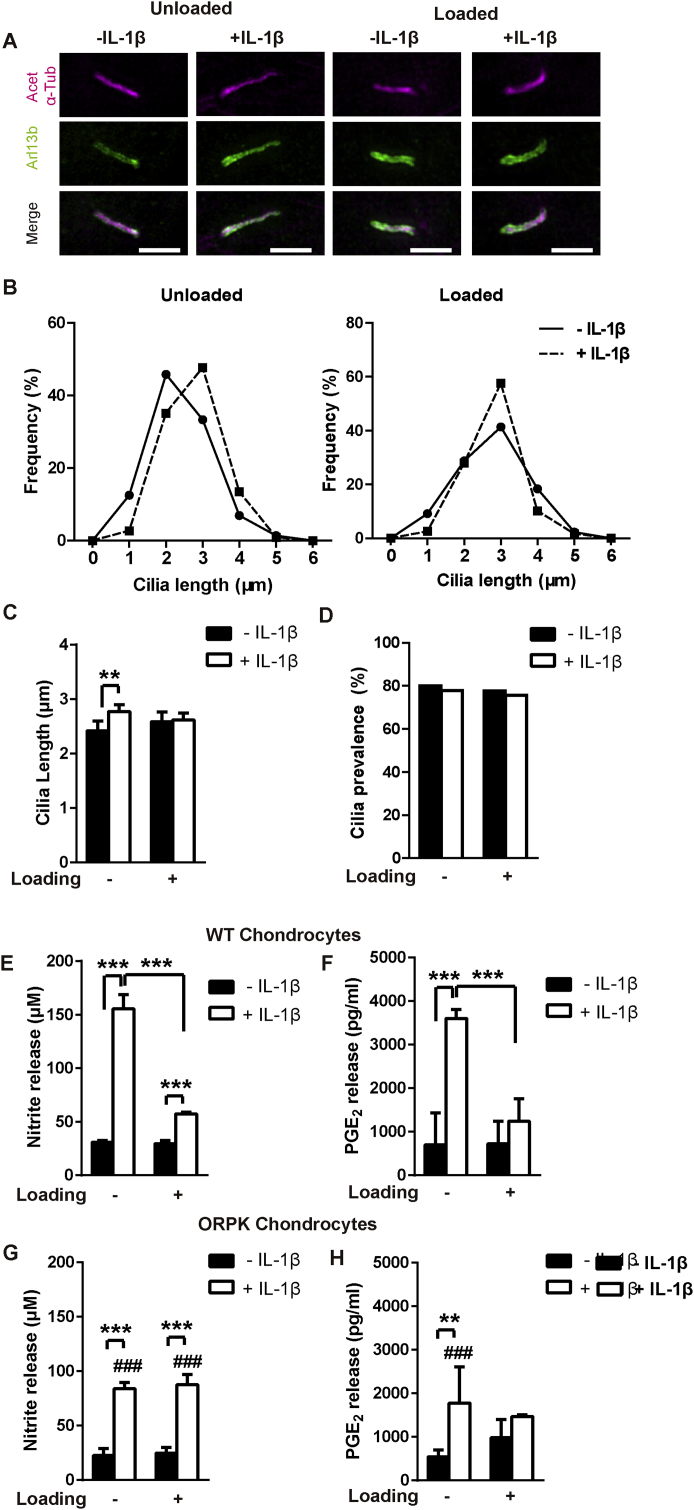
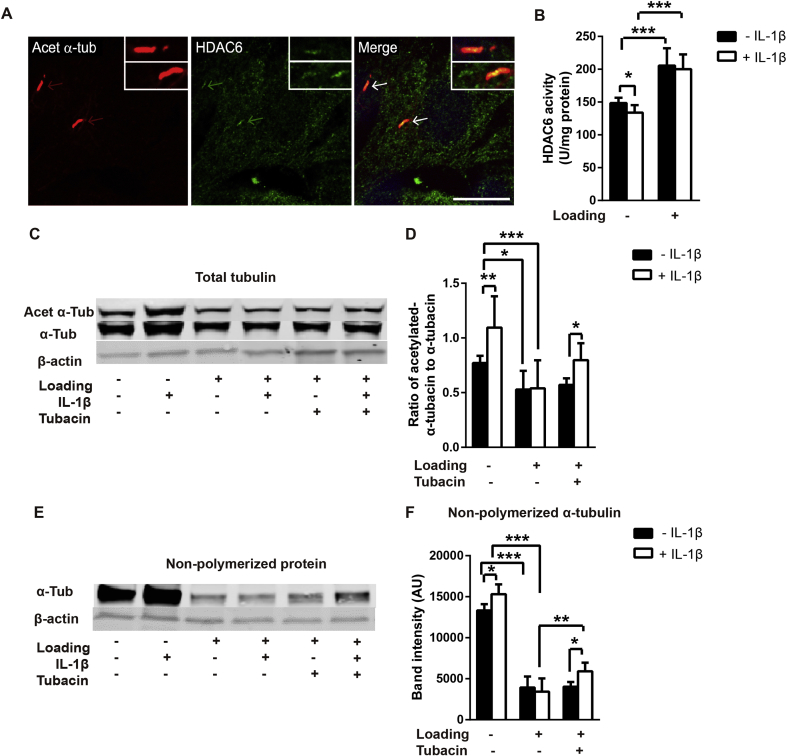
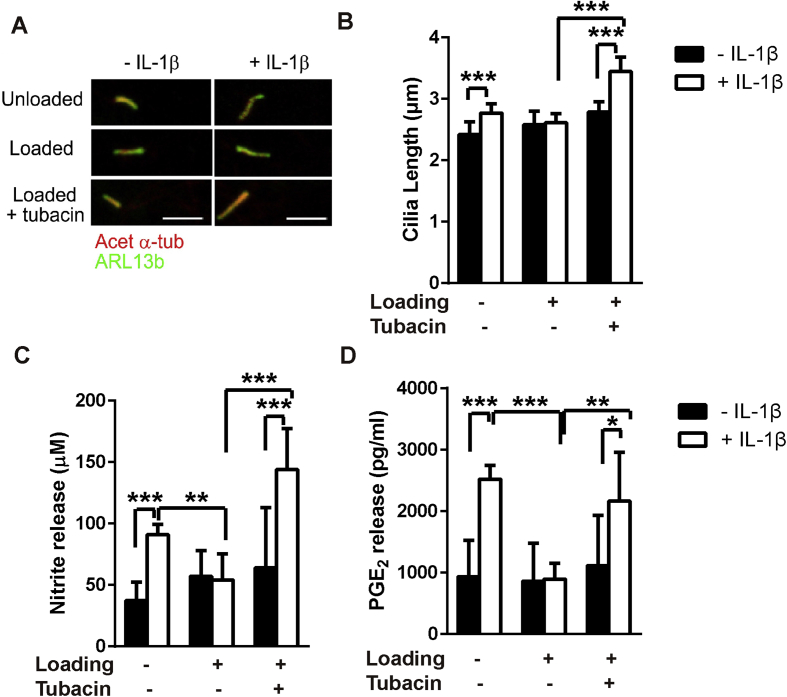
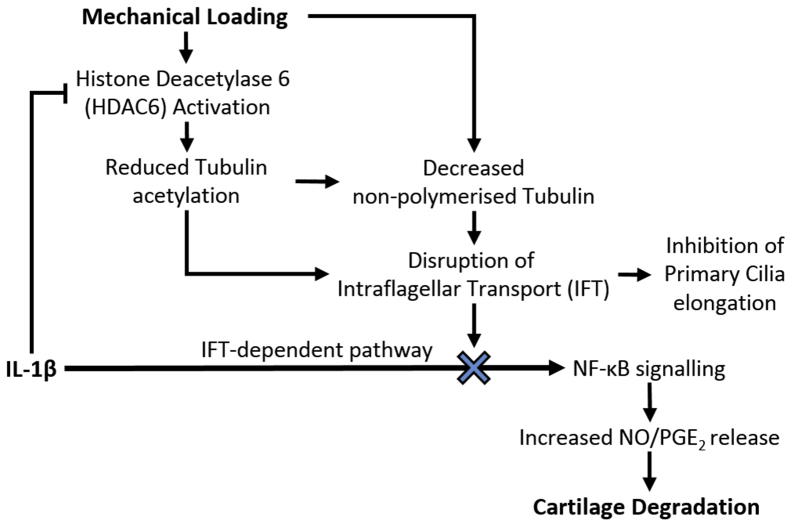
Results: Mechanical loading suppressed NO and PGE2 release and prevented cartilage degradation. Loading activated HDAC6 and disrupted tubulin acetylation and cilia elongation induced by IL-1β. HDAC6 inhibition with tubacin blocked the anti-inflammatory effects of loading and restored tubulin acetylation and cilia elongation. Hypomorphic mutation of IFT88 reduced IL-1β signalling and abolished the anti-inflammatory effects of loading indicating the mechanism is IFT-dependent. Loading reduced the pool of non-polymerised tubulin which was replicated by taxol which also mimicked the anti-inflammatory effects of mechanical loading and prevented cilia elongation.
Conclusions: This study reveals that mechanical loading suppresses inflammatory signalling, partially dependent on IFT, by activation of HDAC6 and post transcriptional modulation of tubulin.
Keywords: Chondrocyte; HDAC6; IFT; IL-1β; Primary cilia; Tubulin.
Copyright © 2019 The Authors. Published by Elsevier Ltd.. All rights reserved.
Figures



References
-
- Singla V., Reiter J.F. The primary cilium as the cell's antenna: signaling at a sensory organelle. Science. 2006;313:629–633. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
