

T cell stemness and dysfunction in tumors are triggered by a common mechanism
- PMID: 30923193
- PMCID: PMC8194369
- DOI: 10.1126/science.aau0135
T cell stemness and dysfunction in tumors are triggered by a common mechanism
Abstract
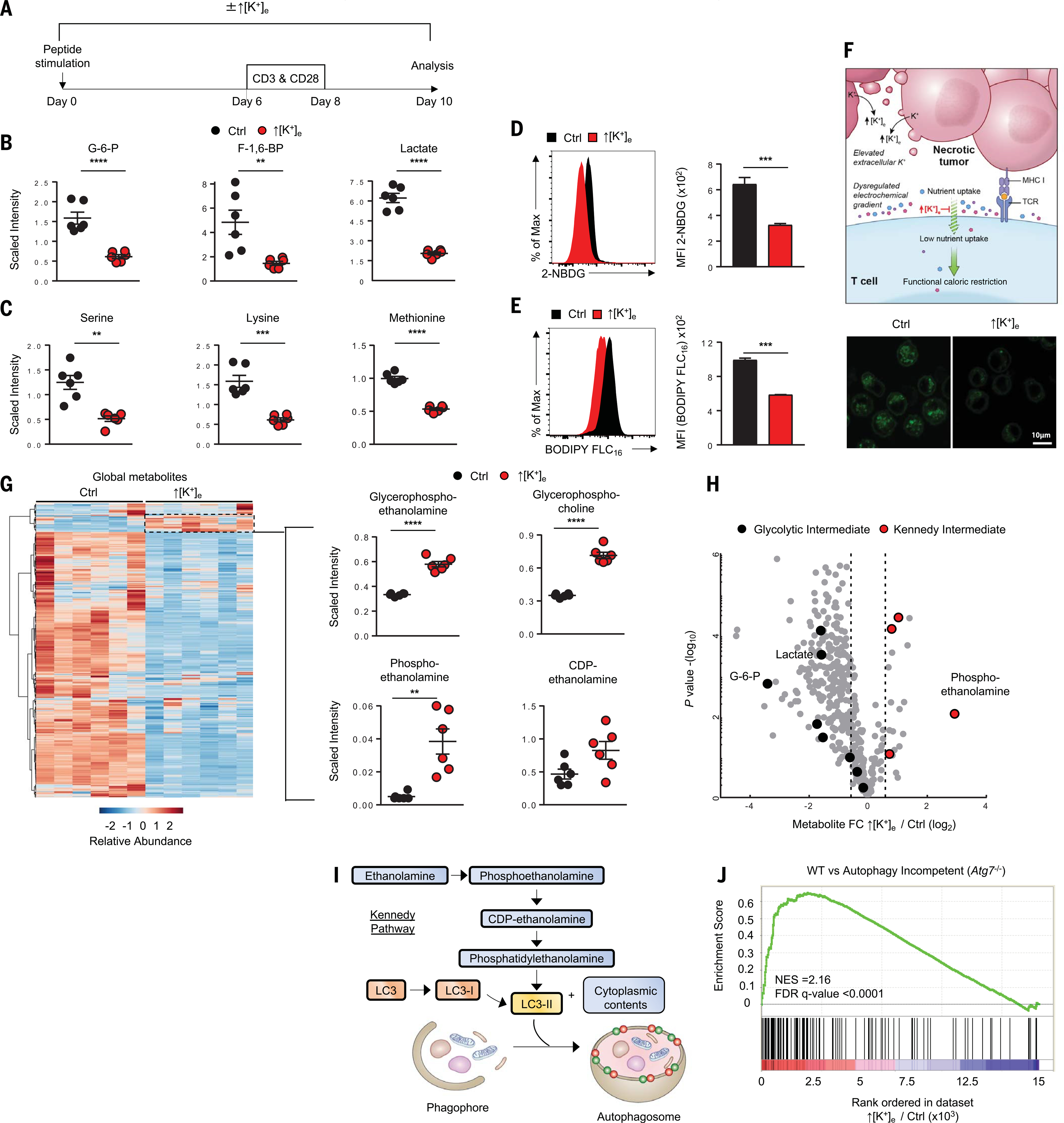
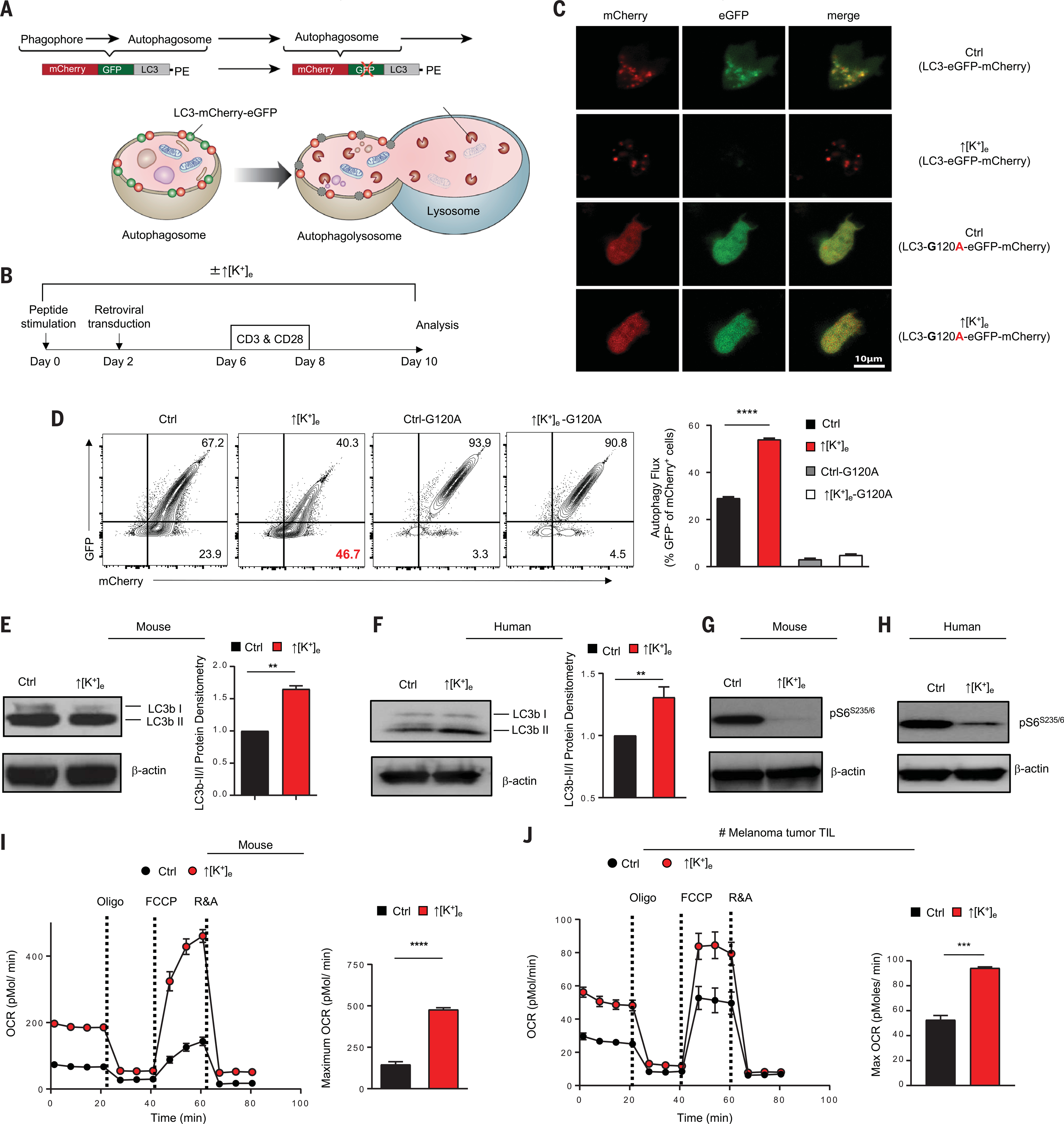
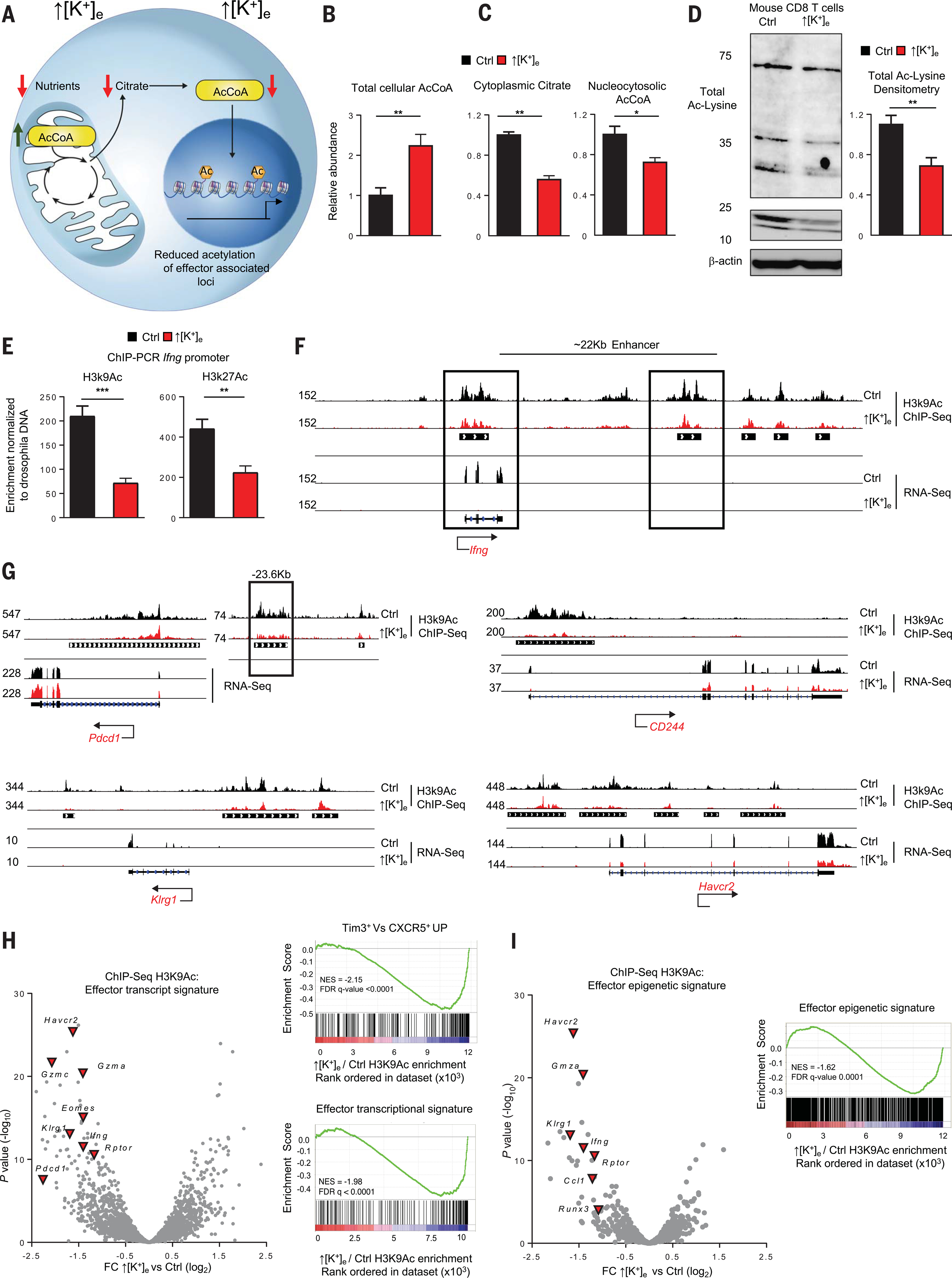
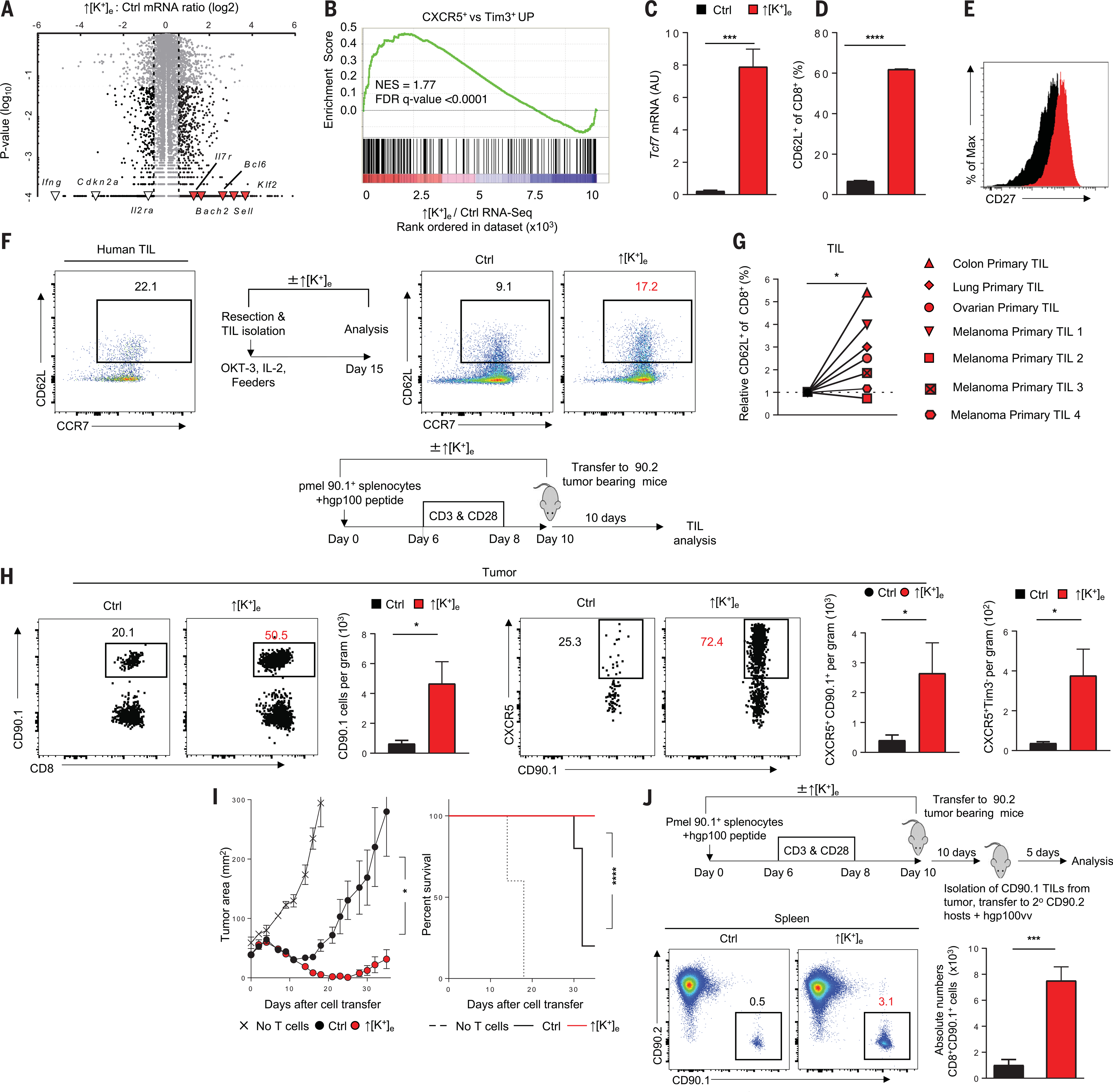
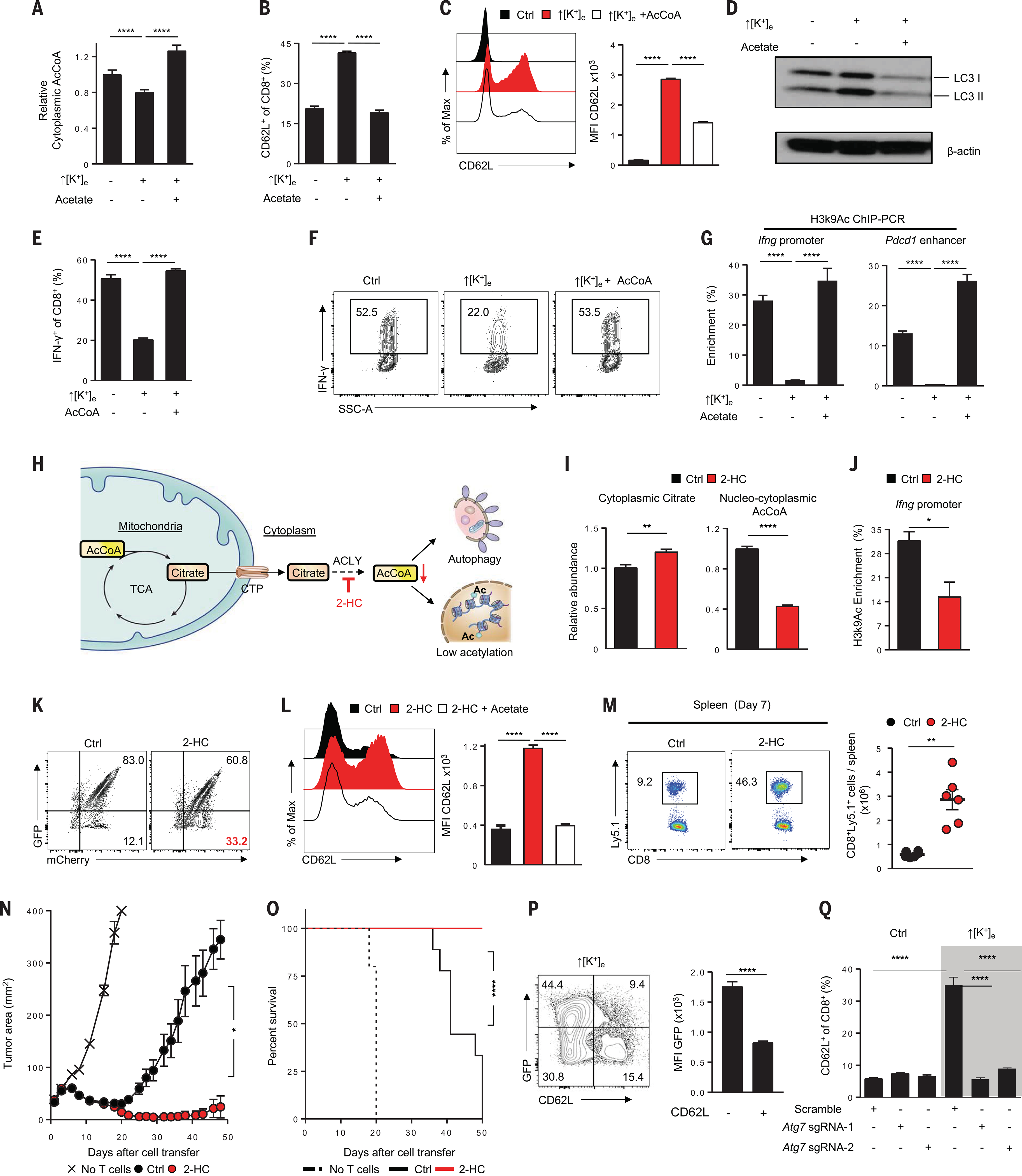
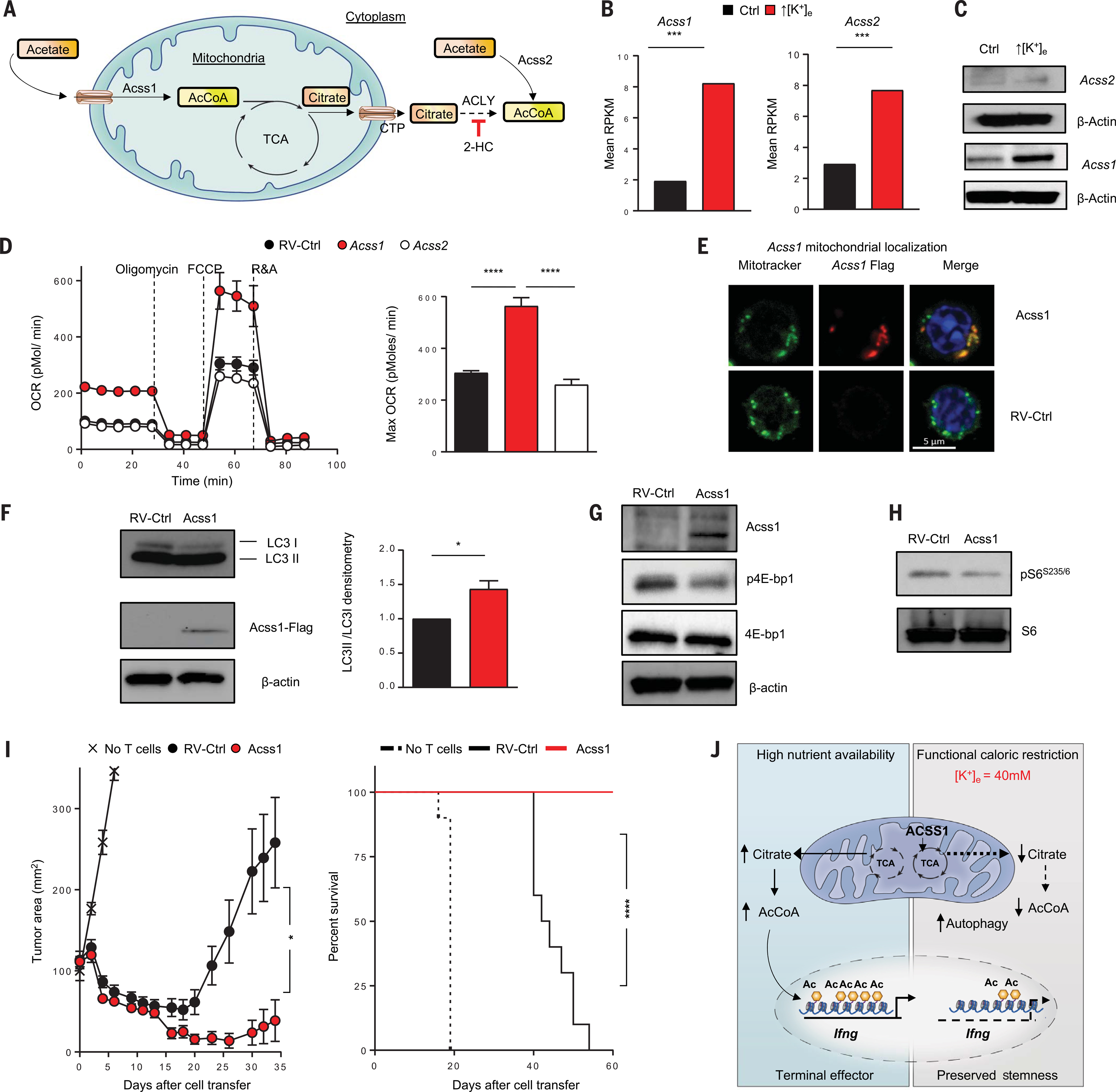
A paradox of tumor immunology is that tumor-infiltrating lymphocytes are dysfunctional in situ, yet are capable of stem cell-like behavior including self-renewal, expansion, and multipotency, resulting in the eradication of large metastatic tumors. We find that the overabundance of potassium in the tumor microenvironment underlies this dichotomy, triggering suppression of T cell effector function while preserving stemness. High levels of extracellular potassium constrain T cell effector programs by limiting nutrient uptake, thereby inducing autophagy and reduction of histone acetylation at effector and exhaustion loci, which in turn produces CD8+ T cells with improved in vivo persistence, multipotency, and tumor clearance. This mechanistic knowledge advances our understanding of T cell dysfunction and may lead to novel approaches that enable the development of enhanced T cell strategies for cancer immunotherapy.
Copyright © 2019 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
Comment in
-
Potassium shapes antitumor immunity.Science. 2019 Mar 29;363(6434):1395-1396. doi: 10.1126/science.aaw8800. Science. 2019. PMID: 30923211 No abstract available.
-
Putting the K+ in K+aloric Restriction.Immunity. 2019 May 21;50(5):1129-1131. doi: 10.1016/j.immuni.2019.04.016. Immunity. 2019. PMID: 31117009
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
