

The hydrogen bonding network of coproheme in coproheme decarboxylase from Listeria monocytogenes: Effect on structure and catalysis
- PMID: 30925402
- PMCID: PMC6517287
- DOI: 10.1016/j.jinorgbio.2019.03.009
The hydrogen bonding network of coproheme in coproheme decarboxylase from Listeria monocytogenes: Effect on structure and catalysis
Abstract
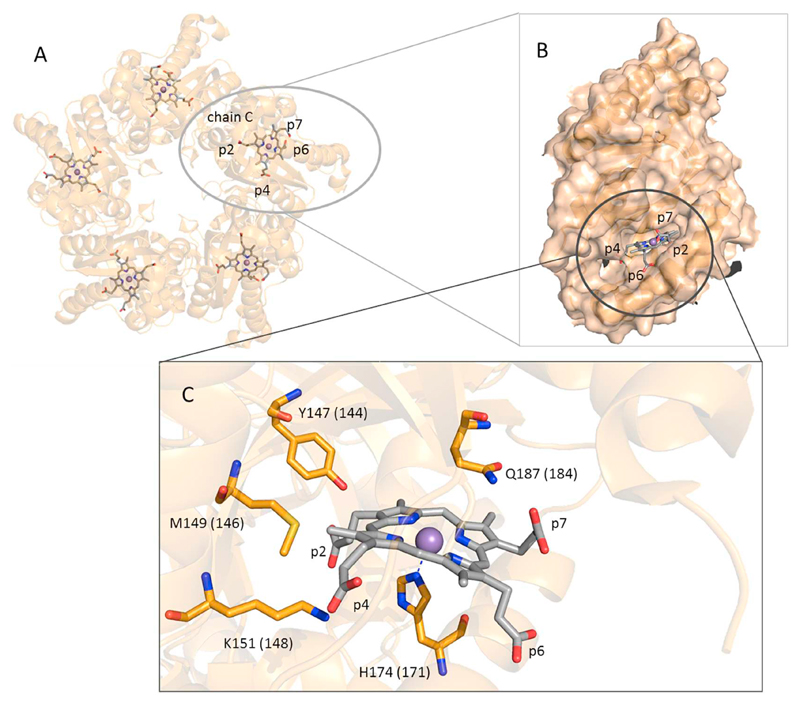
Coproheme decarboxylase (ChdC) catalyzes the oxidative decarboxylation of coproheme to heme b, i.e. the last step in the recently described coproporphyrin-dependent pathway. Coproheme decarboxylation from Listeria monocytogenes is a robust enzymatic reaction of low catalytic efficiency. Coproheme acts as both substrate and redox cofactor activated by H2O2. It fully depends on the catalytic Y147 close to the propionyl group at position 2. In the present study we have investigated the effect of disruption of the comprehensive and conserved hydrogen bonding network between the four propionates and heme cavity residues on (i) the conformational stability of the heme cavity, (ii) the electronic configuration of the ferric redox cofactor/substrate, (iii) the binding of carbon monoxide and, (iv) the decarboxylation reaction mediated by addition of H2O2. Nine single, double and triple mutants of ChdC from Listeria monocytogenes were produced in E. coli. The respective coproheme- and heme b-complexed proteins were studied by UV-Vis, resonance Raman, circular dichroism spectroscopy, and mass spectrometry. Interactions of propionates 2 and 4 with residues in the hydrophobic cavity are crucial for maintenance of the heme cavity architecture, for the mobile distal glutamine to interact with carbon monoxide, and to keep the heme cavity in a closed conformation during turnover. By contrast, the impact of substitution of residues interacting with solvent exposed propionates 6 and 7 was negligible. Except for Y147A and K151A all mutant ChdCs exhibited a wild-type-like catalytic activity. The findings are discussed with respect to the structure-function relationships of ChdCs.
Keywords: Carbon monoxide; Coproheme decarboxylase; Heme b biosynthesis; Propionyl hydrogen-bond; Resonance Raman spectroscopy.
Copyright © 2019 The Authors. Published by Elsevier Inc. All rights reserved.
Figures







References
-
- Lobo SA, Scott A, Videira MA, Winpenny D, Gardner M, Palmer MJ, Schroeder S, Lawrence AD, Parkinson T, Warren MJ, Saraiva LM. Mol Microbiol. 2015;97:472–487. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
