

Loss of postnatal quiescence of neural stem cells through mTOR activation upon genetic removal of cysteine string protein-α
- PMID: 30926666
- PMCID: PMC6475374
- DOI: 10.1073/pnas.1817183116
Loss of postnatal quiescence of neural stem cells through mTOR activation upon genetic removal of cysteine string protein-α
Abstract
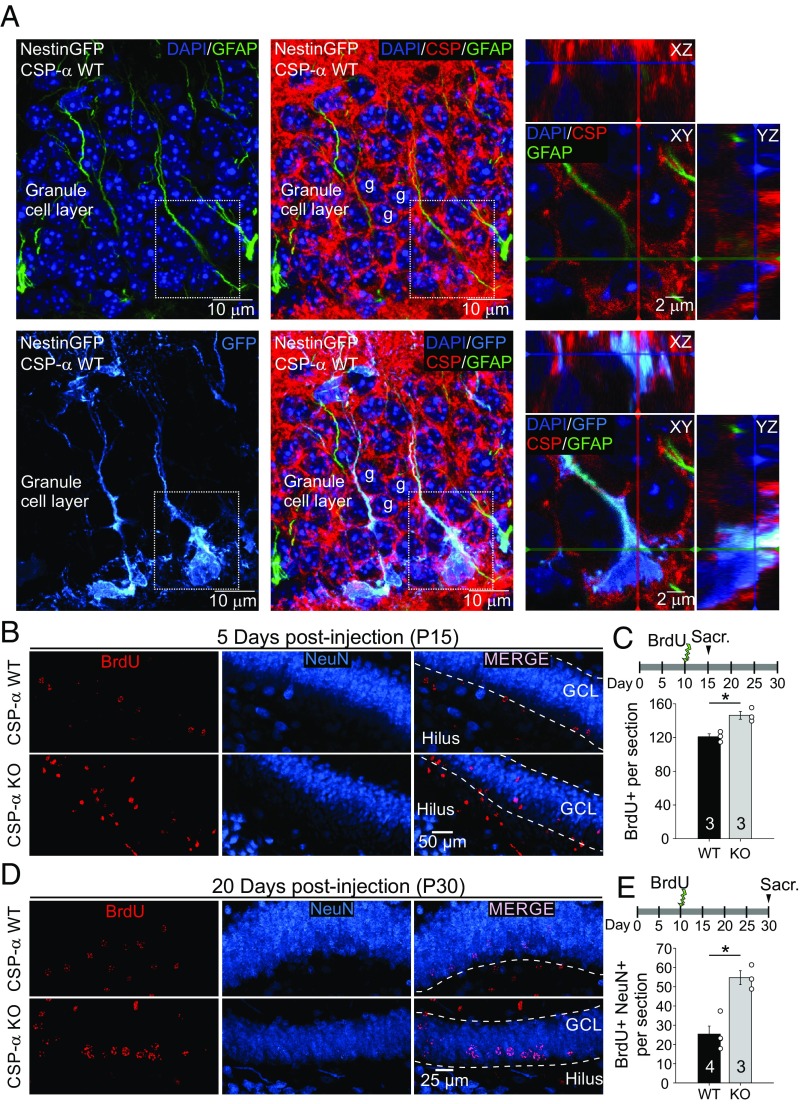
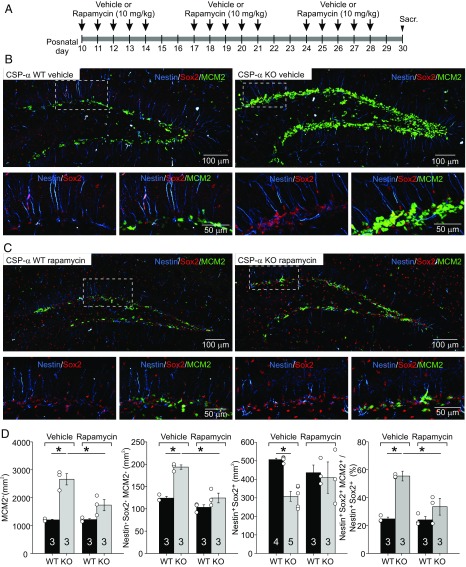
Neural stem cells continuously generate newborn neurons that integrate into and modify neural circuitry in the adult hippocampus. The molecular mechanisms that regulate or perturb neural stem cell proliferation and differentiation, however, remain poorly understood. Here, we have found that mouse hippocampal radial glia-like (RGL) neural stem cells express the synaptic cochaperone cysteine string protein-α (CSP-α). Remarkably, in CSP-α knockout mice, RGL stem cells lose quiescence postnatally and enter into a high-proliferation regime that increases the production of neural intermediate progenitor cells, thereby exhausting the hippocampal neural stem cell pool. In cell culture, stem cells in hippocampal neurospheres display alterations in proliferation for which hyperactivation of the mechanistic target of rapamycin (mTOR) signaling pathway is the primary cause of neurogenesis deregulation in the absence of CSP-α. In addition, RGL cells lose quiescence upon specific conditional targeting of CSP-α in adult neural stem cells. Our findings demonstrate an unanticipated cell-autonomic and circuit-independent disruption of postnatal neurogenesis in the absence of CSP-α and highlight a direct or indirect CSP-α/mTOR signaling interaction that may underlie molecular mechanisms of brain dysfunction and neurodegeneration.
Keywords: DNAJC5; adult neurogenesis; adult-onset neuronal ceroid lipofuscinosis; lysosome; synaptic neurodegeneration.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
