

Shared Cortex-Cerebellum Dynamics in the Execution and Learning of a Motor Task
- PMID: 30929904
- PMCID: PMC6500577
- DOI: 10.1016/j.cell.2019.02.019
Shared Cortex-Cerebellum Dynamics in the Execution and Learning of a Motor Task
Abstract
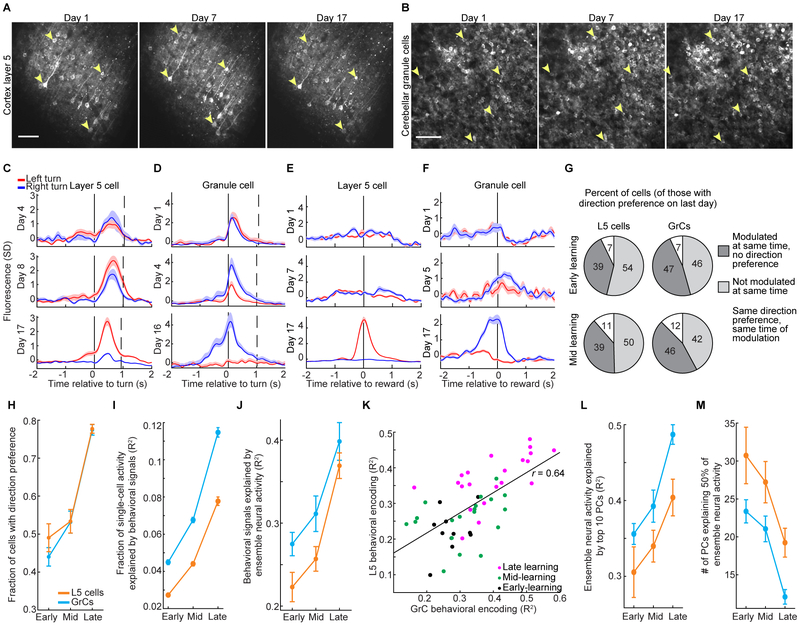
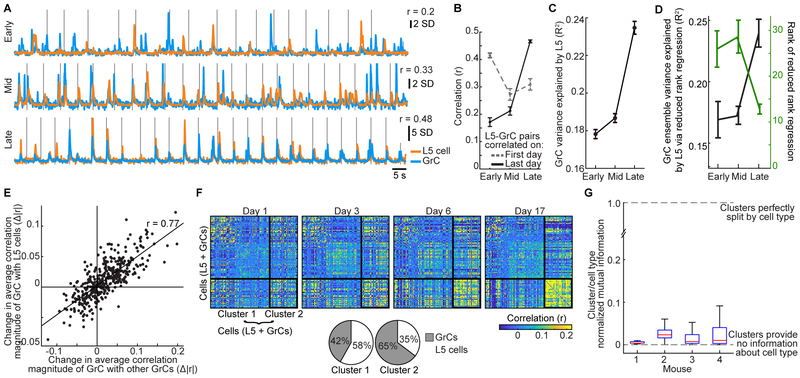
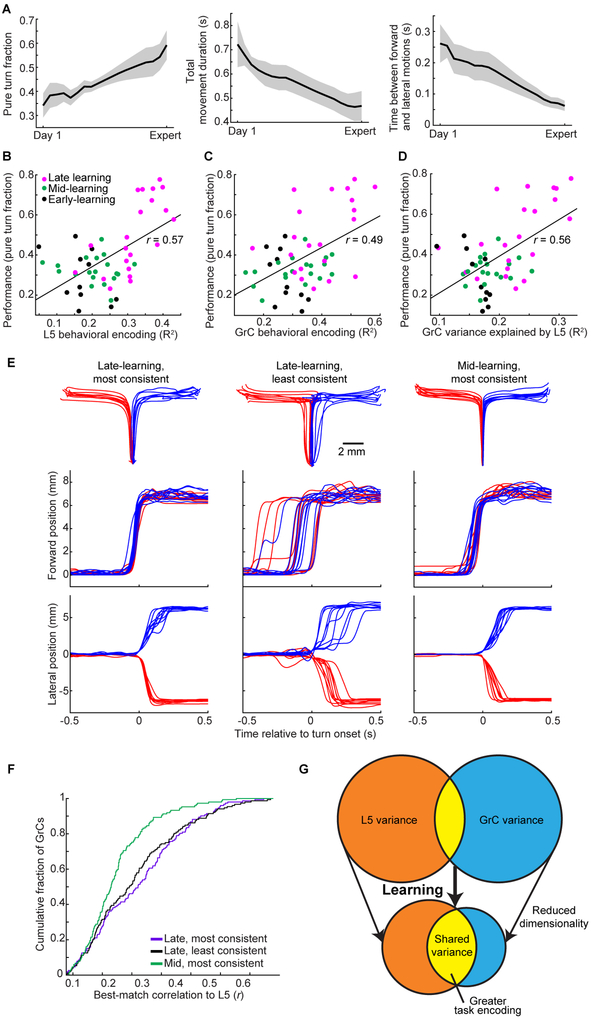
Throughout mammalian neocortex, layer 5 pyramidal (L5) cells project via the pons to a vast number of cerebellar granule cells (GrCs), forming a fundamental pathway. Yet, it is unknown how neuronal dynamics are transformed through the L5→GrC pathway. Here, by directly comparing premotor L5 and GrC activity during a forelimb movement task using dual-site two-photon Ca2+ imaging, we found that in expert mice, L5 and GrC dynamics were highly similar. L5 cells and GrCs shared a common set of task-encoding activity patterns, possessed similar diversity of responses, and exhibited high correlations comparable to local correlations among L5 cells. Chronic imaging revealed that these dynamics co-emerged in cortex and cerebellum over learning: as behavioral performance improved, initially dissimilar L5 cells and GrCs converged onto a shared, low-dimensional, task-encoding set of neural activity patterns. Thus, a key function of cortico-cerebellar communication is the propagation of shared dynamics that emerge during learning.
Keywords: brain state; cerebellum; dimensional expansion; granule cells; layer 5; motor learning; movement planning; neocortex; pontine nuclei; reward.
Copyright © 2019 Elsevier Inc. All rights reserved.
Figures
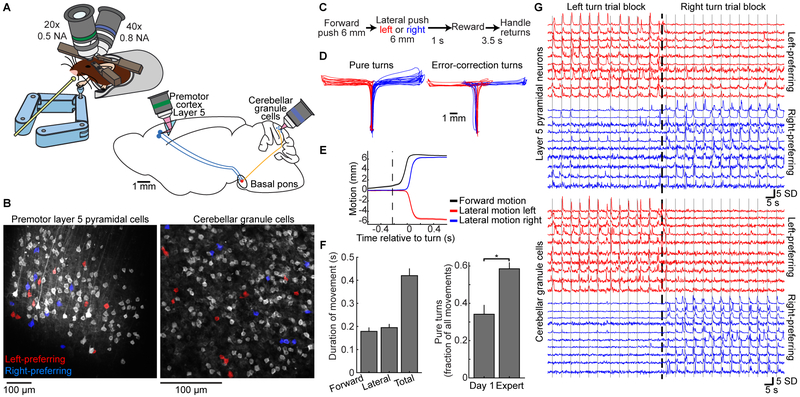
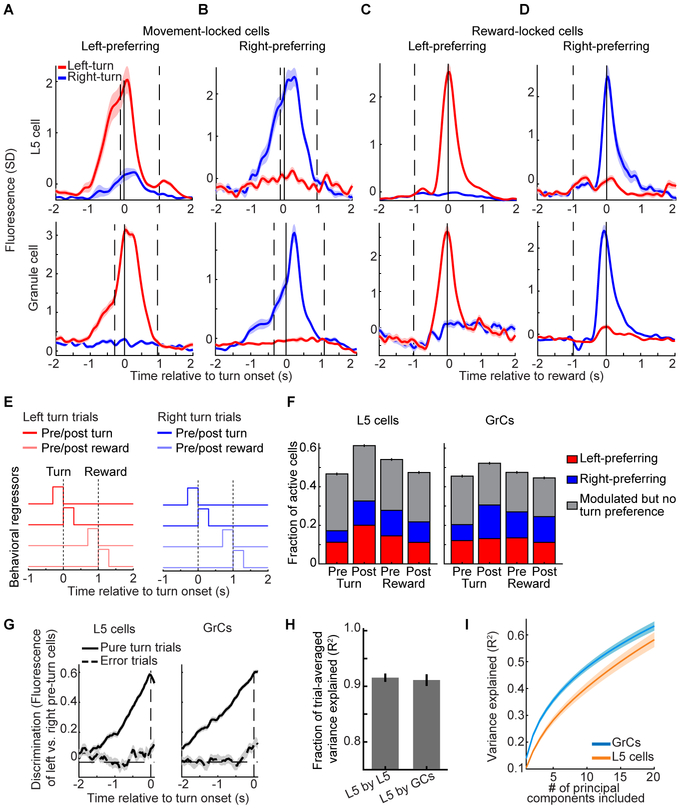
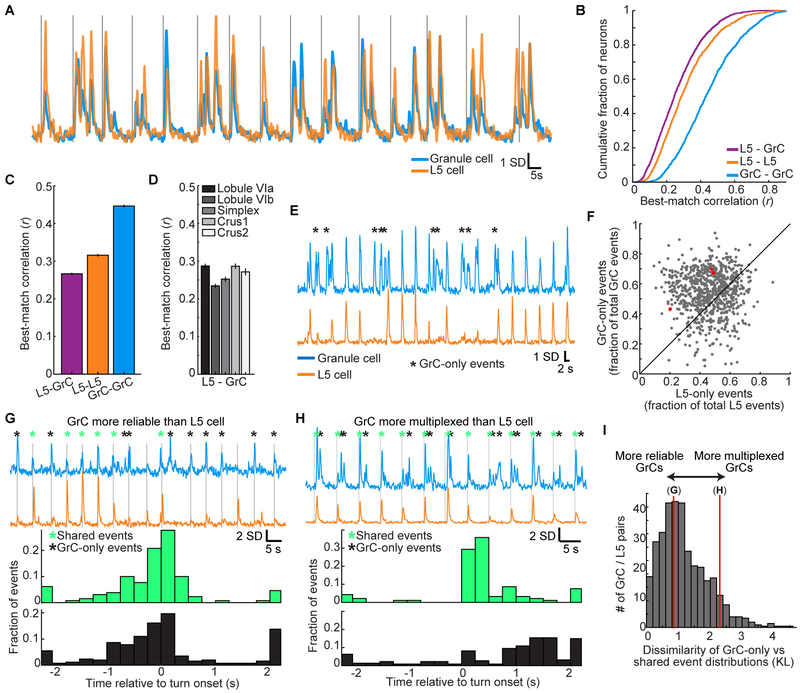
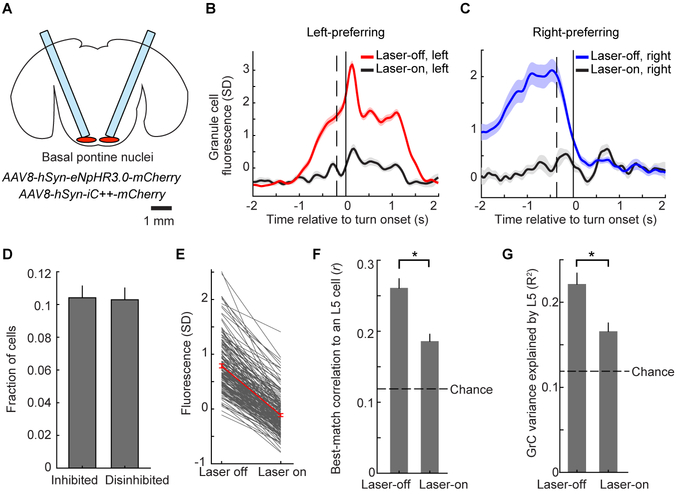
References
-
- Albus JS (1971). A theory of cerebellar function. Mathematical Biosciences 10, 25–61.
-
- Babadi B, and Sompolinsky H (2014). Sparseness and expansion in sensory representations. Neuron 83, 1213–1226. - PubMed
-
- Barton Robert A., and Venditti C (2014). Rapid Evolution of the Cerebellum in Humans and Other Great Apes. Curr Biol 24, 2440–2444. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
