

The Activation State of CD4 T Cells Alters Cellular Peptidase Activities, HIV Antigen Processing, and MHC Class I Presentation in a Sequence-Dependent Manner
- PMID: 30936293
- PMCID: PMC6504579
- DOI: 10.4049/jimmunol.1700950
The Activation State of CD4 T Cells Alters Cellular Peptidase Activities, HIV Antigen Processing, and MHC Class I Presentation in a Sequence-Dependent Manner
Abstract
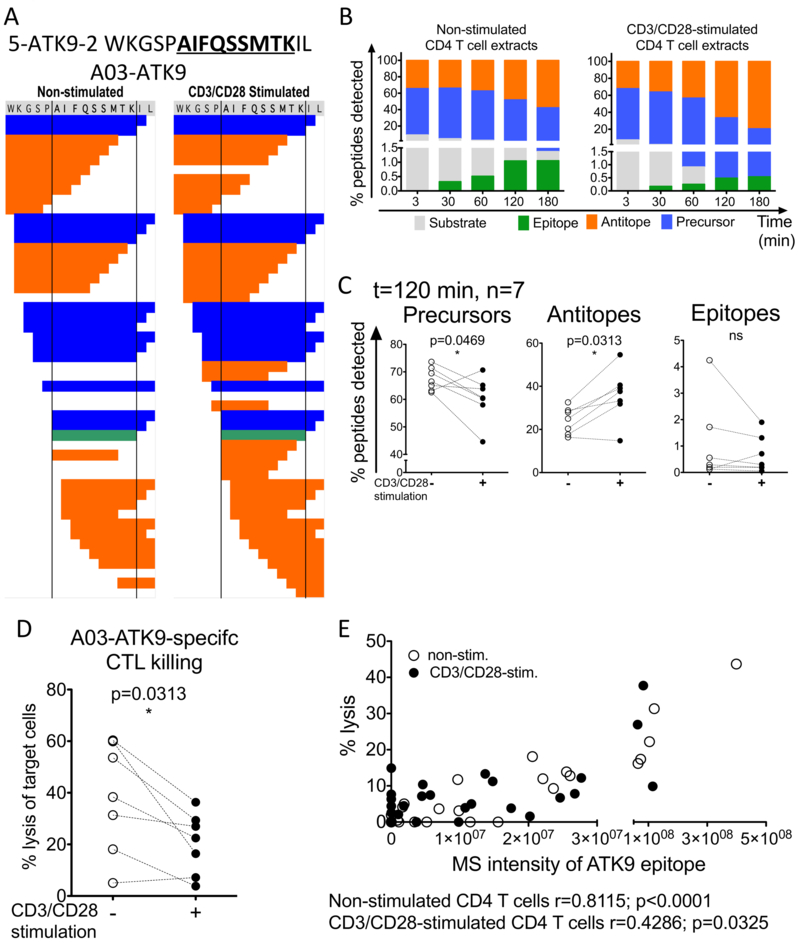
CD4 T cell activation is critical to the initiation of adaptive immunity. CD4 T cells are also the main targets of HIV infection, and their activation status contributes to the maintenance and outcome of infection. Although the role of activation in the differentiation and proliferation of CD4 T cells is well studied, its impact on the processing and MHC class I (MHC-I) presentation of epitopes and immune recognition by CD8 T cells are not investigated. In this study, we show that the expression and hydrolytic activities of cellular peptidases are increased upon TCR-dependent and MHC-peptide activation of primary CD4 T cells from healthy or HIV-infected persons. Changes in peptidase activities altered the degradation patterns of HIV Ags analyzed by mass spectrometry, modifying the amount of MHC-I epitopes produced, the antigenicity of the degradation products, and the coverage of Ags by degradation peptides presentable by MHC-I. The computational analysis of 2237 degradation peptides generated during the degradation of various HIV-antigenic fragments in CD4 T cells identified cleavage sites that were predictably enhanced, reduced, or unchanged upon cellular activation. Epitope processing and presentation by CD4 T cells may be modulated by the activation state of cells in a sequence-dependent manner. Accordingly, cellular activation modified endogenous Ag processing and presentation and killing of HIV-infected CD4 T cells by CD8 T cells in a way that mirrored differences in in vitro epitope processing. The clearance of HIV-infected cells may rely on different immune responses according to activation state during HIV infection.
Copyright © 2019 by The American Association of Immunologists, Inc.
Figures






References
-
- Baldauf HM, Pan X, Erikson E, Schmidt S, Daddacha W, Burggraf M, Schenkova K, Ambiel I, Wabnitz G, Gramberg T, Panitz S, Flory E, Landau NR, Sertel S, Rutsch F, Lasitschka F, Kim B, Konig R, Fackler OT, and Keppler OT. 2012. SAMHD1 restricts HIV-1 infection in resting CD4(+) T cells. Nat Med 18: 1682–1687. - PMC - PubMed
-
- Ruffin N, Brezar V, Ayinde D, Lefebvre C, Schulze Zur Wiesch J, van Lunzen J, Bockhorn M, Schwartz O, Hocini H, Lelievre JD, Banchereau J, Levy Y, and Seddiki N. 2015. Low SAMHD1 expression following T-cell activation and proliferation renders CD4+ T cells susceptible to HIV-1. AIDS 29: 519–530. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
