

Epigenetic Priming in Drug Addiction
- PMID: 30936392
- PMCID: PMC6764605
- DOI: 10.1101/sqb.2018.83.037663
Epigenetic Priming in Drug Addiction
Abstract
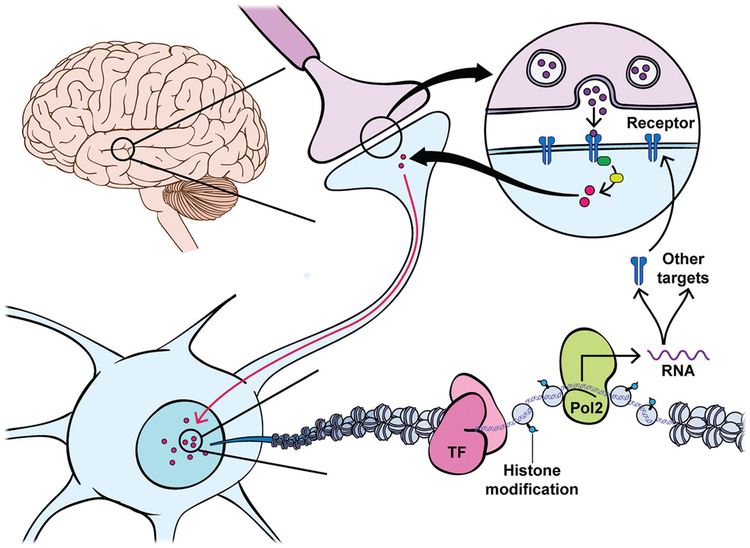
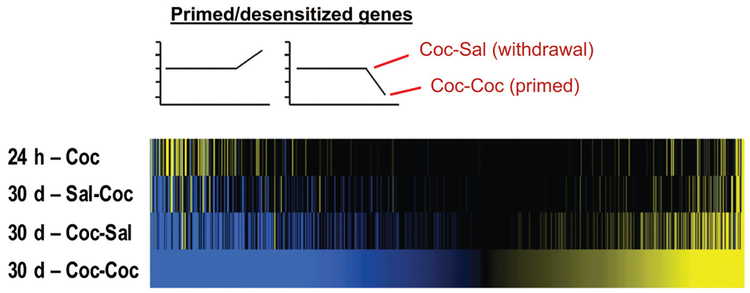
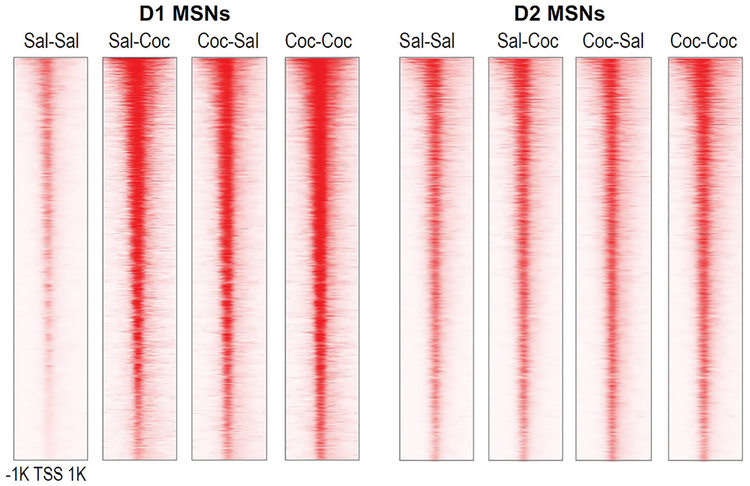
Drug addiction is a chronic relapsing brain disorder that is characterized by compulsive drug seeking and continued use despite negative outcomes. Current pharmacological therapies target neuronal receptors or transporters upon which drugs of abuse act initially, yet these treatments remain ineffective for most individuals and do not prevent disease relapse after abstinence. Drugs of abuse, in addition to their acute effects, cause persistent plasticity after repeated use, involving dysregulated gene expression in the brain's reward regions, which are thought to mediate the persistent behavioral abnormalities that characterize addiction. Emerging evidence implicates epigenetic priming as a key mechanism that underlies the long-lasting alterations in neuronal gene regulation, which can remain latent until triggered by re-exposure to drug-associated stimuli or the drug itself. Thus, to effectively treat drug addiction, we must identify the precise epigenetic mechanisms that establish and preserve the drug-induced pathology of the brain reward circuitry.
© 2018 Mews et al.; Published by Cold Spring Harbor Laboratory Press.
Figures

References
Grants and funding
LinkOut - more resources
Full Text Sources