

Expectation-induced modulation of metastable activity underlies faster coding of sensory stimuli
- PMID: 30936557
- PMCID: PMC6516078
- DOI: 10.1038/s41593-019-0364-9
Expectation-induced modulation of metastable activity underlies faster coding of sensory stimuli
Abstract
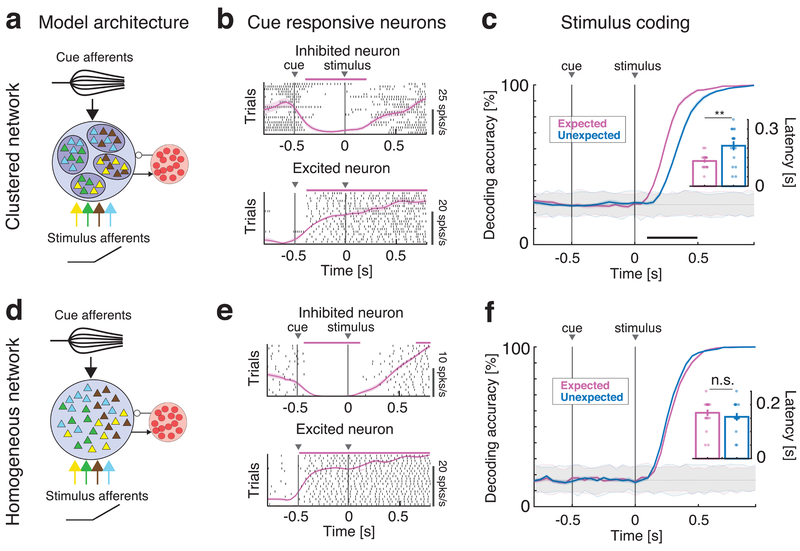
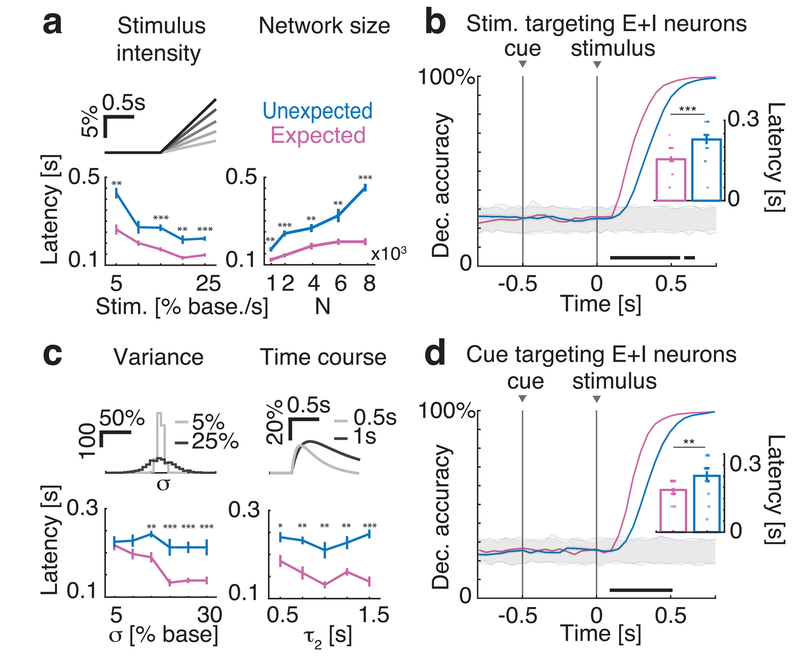
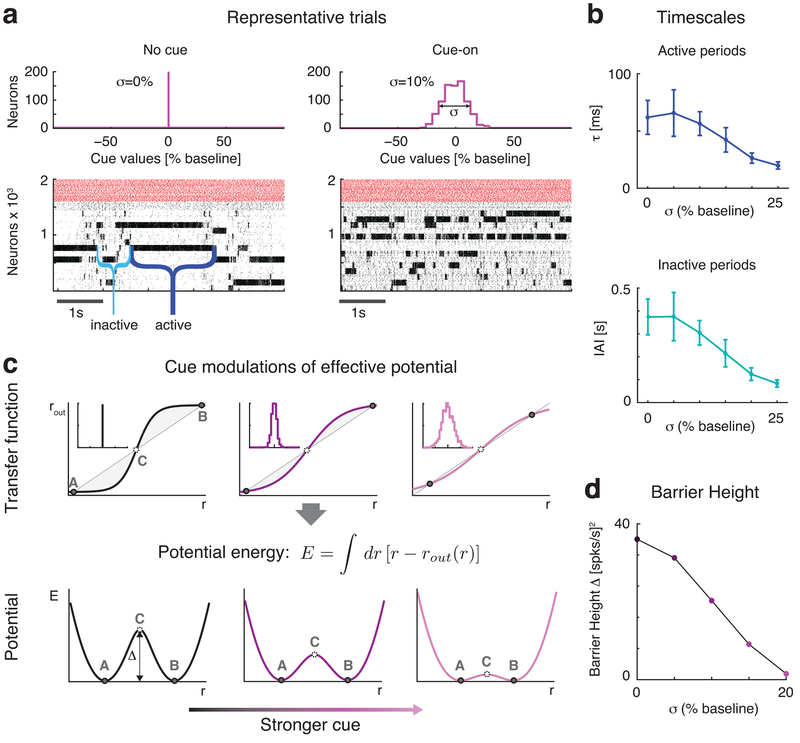
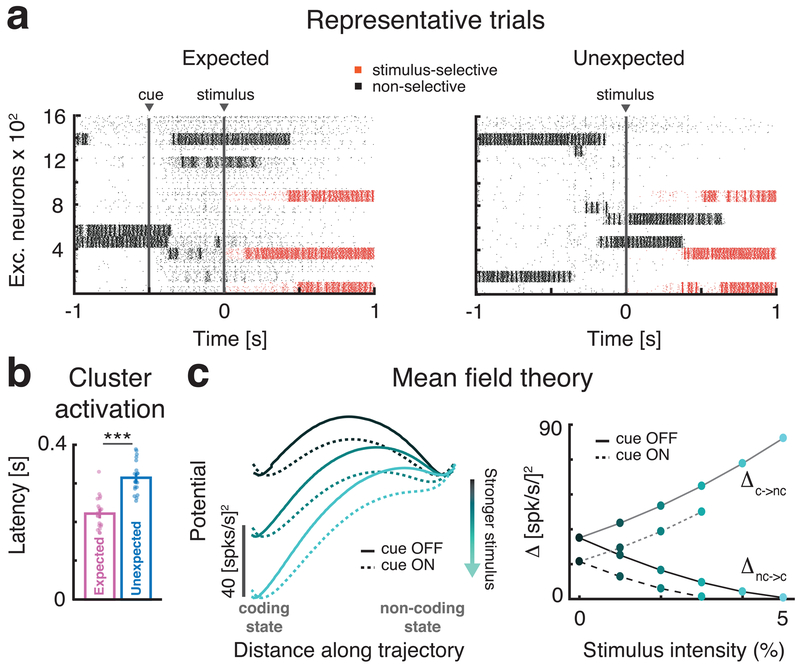
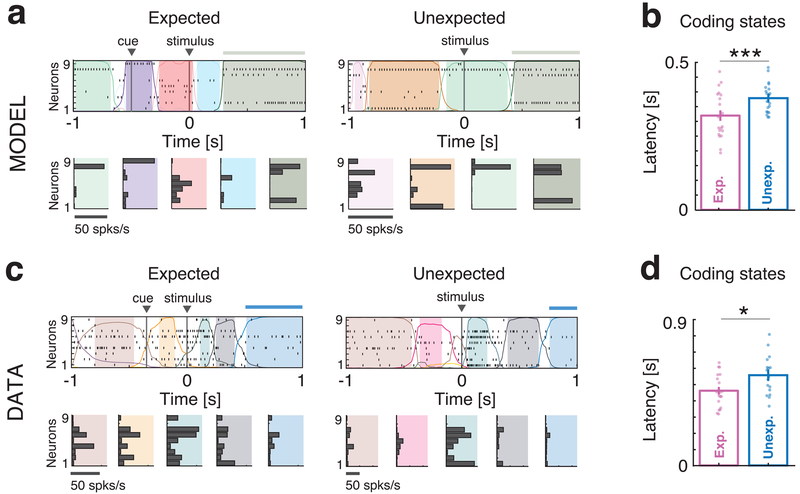
Sensory stimuli can be recognized more rapidly when they are expected. This phenomenon depends on expectation affecting the cortical processing of sensory information. However, the mechanisms responsible for the effects of expectation on sensory circuits remain elusive. In the present study, we report a novel computational mechanism underlying the expectation-dependent acceleration of coding observed in the gustatory cortex of alert rats. We use a recurrent spiking network model with a clustered architecture capturing essential features of cortical activity, such as its intrinsically generated metastable dynamics. Relying on network theory and computer simulations, we propose that expectation exerts its function by modulating the intrinsically generated dynamics preceding taste delivery. Our model's predictions were confirmed in the experimental data, demonstrating how the modulation of ongoing activity can shape sensory coding. Altogether, these results provide a biologically plausible theory of expectation and ascribe an alternative functional role to intrinsically generated, metastable activity.
Conflict of interest statement
Competing Financial Interests
The authors declare no competing financial interests.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
