

The Endoplasmic Reticulum-Mitochondria Encounter Structure Complex Coordinates Coenzyme Q Biosynthesis
- PMID: 30937424
- PMCID: PMC6441334
- DOI: 10.1177/2515256418825409
The Endoplasmic Reticulum-Mitochondria Encounter Structure Complex Coordinates Coenzyme Q Biosynthesis
Abstract
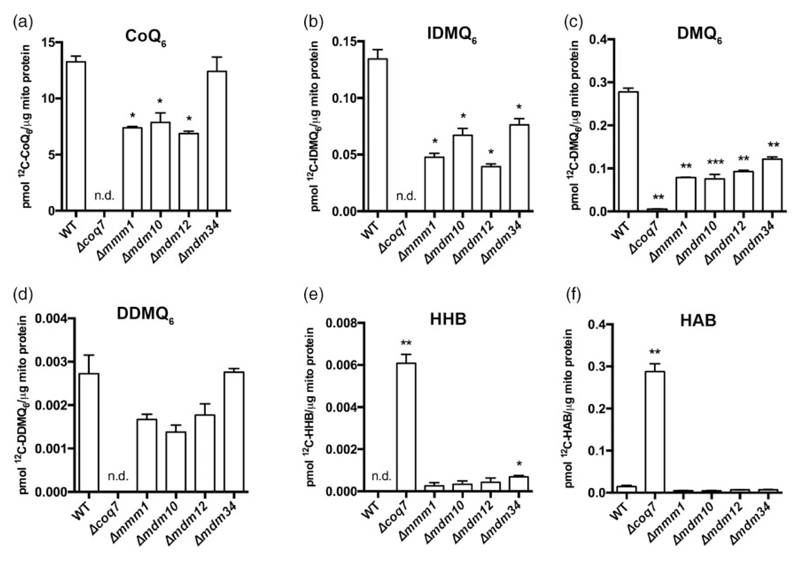
Loss of the endoplasmic reticulum (ER)-mitochondria encounter structure (ERMES) complex that resides in contact sites between the yeast ER and mitochondria leads to impaired respiration; however, the reason for that is not clear. We find that in ERMES null mutants, there is an increase in the level of mRNAs encoding for biosynthetic enzymes of coenzyme Q6 (CoQ6), an essential electron carrier of the mitochondrial respiratory chain. We show that the mega complexes involved in CoQ6 biosynthesis (CoQ synthomes) are destabilized in ERMES mutants. This, in turn, affects the level and distribution of CoQ6 within the cell, resulting in reduced mitochondrial CoQ6. We suggest that these outcomes contribute to the reduced respiration observed in ERMES mutants. Fluorescence microscopy experiments demonstrate close proximity between the CoQ synthome and ERMES, suggesting a spatial coordination. The involvement of the ER-mitochondria contact site in regulation of CoQ6 biogenesis highlights an additional level of communication between these two organelles.
Keywords: ER-mitochondrial encounter structure; coenzyme Q; endoplasmic reticulum; mitochondrion (mitochondria).
Conflict of interest statement
Declaration of Conflicting Interests The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Figures




References
-
- Allan CM, Hill S, Morvaridi S, Saiki R, Johnson JS, Liau WS, Clarke CF. A conserved START domain coenzyme Q-binding polypeptide is required for efficient Q biosynthesis, respiratory electron transport, and antioxidant function in Saccharomyces cerevisiae. Biochimica et Biophysica Acta – Molecular and Cell Biology of Lipids. 2013;1831:776–791. - PMC - PubMed
-
- Barros MH, Johnson A, Gin P, Marbois BN, Clarke CF, Tzagoloff A. The Saccharomyces cerevisiae COQ10 gene encodes a START domain protein required for function of coenzyme Q in respiration. Journal of Biological Chemistry. 2005;280:42627–42635. - PubMed
-
- Belogrudov GI, Lee PT, Jonassen T, Hsu AY, Gin P, Clarke CF. Yeast COQ4 encodes a mitochondrial protein required for coenzyme Q synthesis. Archives of Biochemistry and Biophysics. 2001;392:48–58. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases