

Identification of a Novel Deltavirus in Boa Constrictors
- PMID: 30940697
- PMCID: PMC6445931
- DOI: 10.1128/mBio.00014-19
Identification of a Novel Deltavirus in Boa Constrictors
Abstract
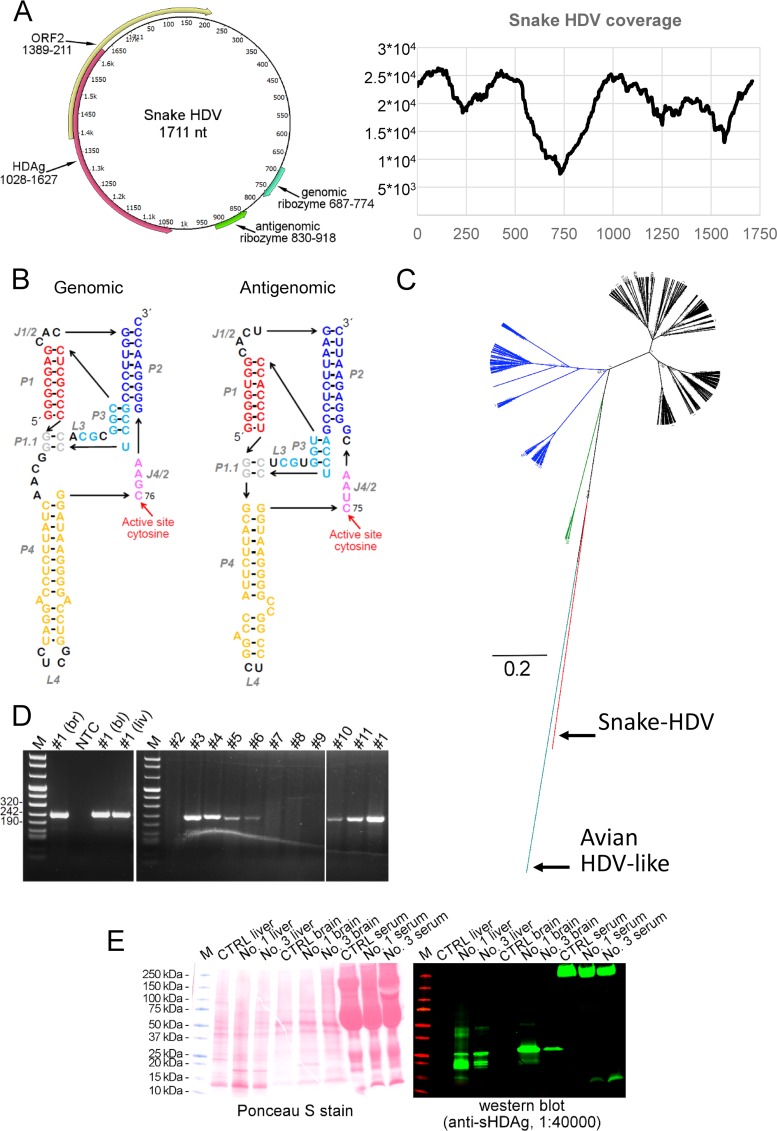
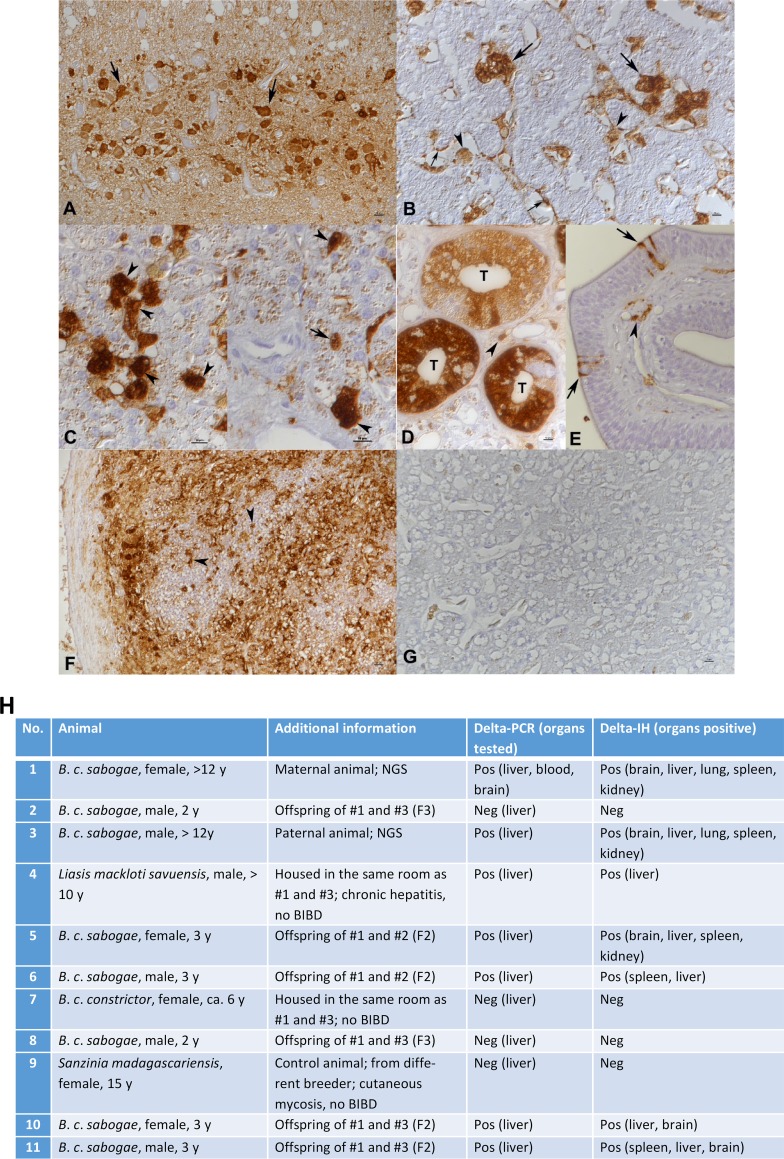
Hepatitis D virus (HDV) forms the genus Deltavirus unassigned to any virus family. HDV is a satellite virus and needs hepatitis B virus (HBV) to make infectious particles. Deltaviruses are thought to have evolved in humans, since for a long time, they had not been identified elsewhere. Herein we report, prompted by the recent discovery of an HDV-like agent in birds, the identification of a deltavirus in snakes (Boa constrictor) designated snake HDV (sHDV). The circular 1,711-nt RNA genome of sHDV resembles human HDV (hHDV) in its coding strategy and size. We discovered sHDV during a metatranscriptomic study of brain samples of a Boa constrictor breeding pair with central nervous system signs. Applying next-generation sequencing (NGS) to brain, blood, and liver samples from both snakes, we did not find reads matching hepadnaviruses. Sequence comparison showed the snake delta antigen (sHDAg) to be 55% and 37% identical to its human and avian counterparts. Antiserum raised against recombinant sHDAg was used in immunohistology and demonstrated a broad viral target cell spectrum, including neurons, epithelial cells, and leukocytes. Using RT-PCR, we also detected sHDV RNA in two juvenile offspring and in a water python (Liasis macklotisavuensis) in the same snake colony, potentially indicating vertical and horizontal transmission. Screening of 20 randomly selected boas from another breeder by RT-PCR revealed sHDV infection in three additional snakes. The observed broad tissue tropism and the failure to detect accompanying hepadnavirus suggest that sHDV could be a satellite virus of a currently unknown enveloped virus.IMPORTANCE So far, the only known example of deltaviruses is the hepatitis delta virus (HDV). HDV is speculated to have evolved in humans, since deltaviruses were until very recently found only in humans. Using a metatranscriptomic sequencing approach, we found a circular RNA, which resembles that of HDV in size and coding strategy, in a snake. The identification of similar deltaviruses in distantly related species other than humans indicates that the previously suggested hypotheses on the origins of deltaviruses need to be updated. It is still possible that the ancestor of deltaviruses emerged from cellular RNAs; however, it likely would have happened much earlier in evolution than previously thought. These findings open up completely new avenues in evolution and pathogenesis studies of deltaviruses.
Keywords: deltavirus; evolutionary biology; hepatitis; virology; zoonotic infections.
Copyright © 2019 Hetzel et al.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
