

T cell-mediated immunity to malaria
- PMID: 30940932
- PMCID: PMC6599480
- DOI: 10.1038/s41577-019-0158-z
T cell-mediated immunity to malaria
Abstract
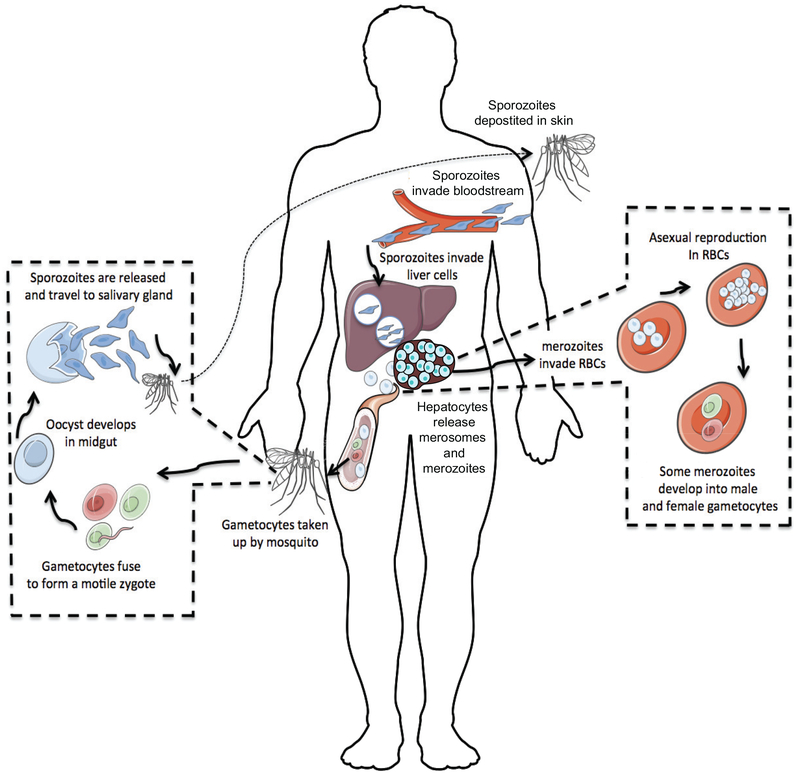
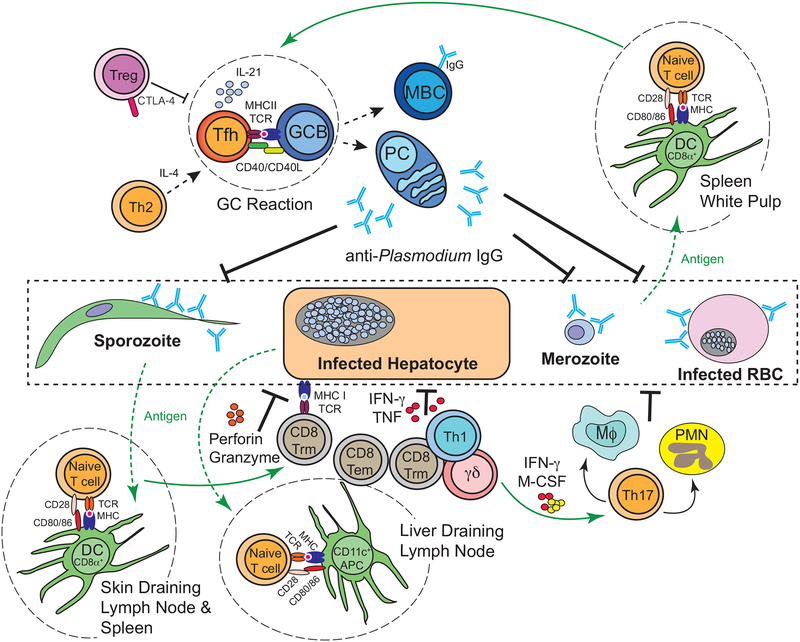
Immunity to malaria has been linked to the availability and function of helper CD4+ T cells, cytotoxic CD8+ T cells and γδ T cells that can respond to both the asymptomatic liver stage and the symptomatic blood stage of Plasmodium sp. infection. These T cell responses are also thought to be modulated by regulatory T cells. However, the precise mechanisms governing the development and function of Plasmodium-specific T cells and their capacity to form tissue-resident and long-lived memory populations are less well understood. The field has arrived at a point where the push for vaccines that exploit T cell-mediated immunity to malaria has made it imperative to define and reconcile the mechanisms that regulate the development and functions of Plasmodium-specific T cells. Here, we review our current understanding of the mechanisms by which T cell subsets orchestrate host resistance to Plasmodium infection on the basis of observational and mechanistic studies in humans, non-human primates and rodent models. We also examine the potential of new experimental strategies and human infection systems to inform a new generation of approaches to harness T cell responses against malaria.
Figures
References
-
- Regules JA et al. Fractional Third and Fourth Dose of RTS,S/AS01 Malaria Candidate Vaccine: A Phase 2a Controlled Human Malaria Parasite Infection and Immunogenicity Study. J Infect Dis 214, 762–771 (2016). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
