

Yb body assembly on the flamenco piRNA precursor transcripts reduces genic piRNA production
- PMID: 30943101
- PMCID: PMC6724695
- DOI: 10.1091/mbc.E17-10-0591
Yb body assembly on the flamenco piRNA precursor transcripts reduces genic piRNA production
Abstract
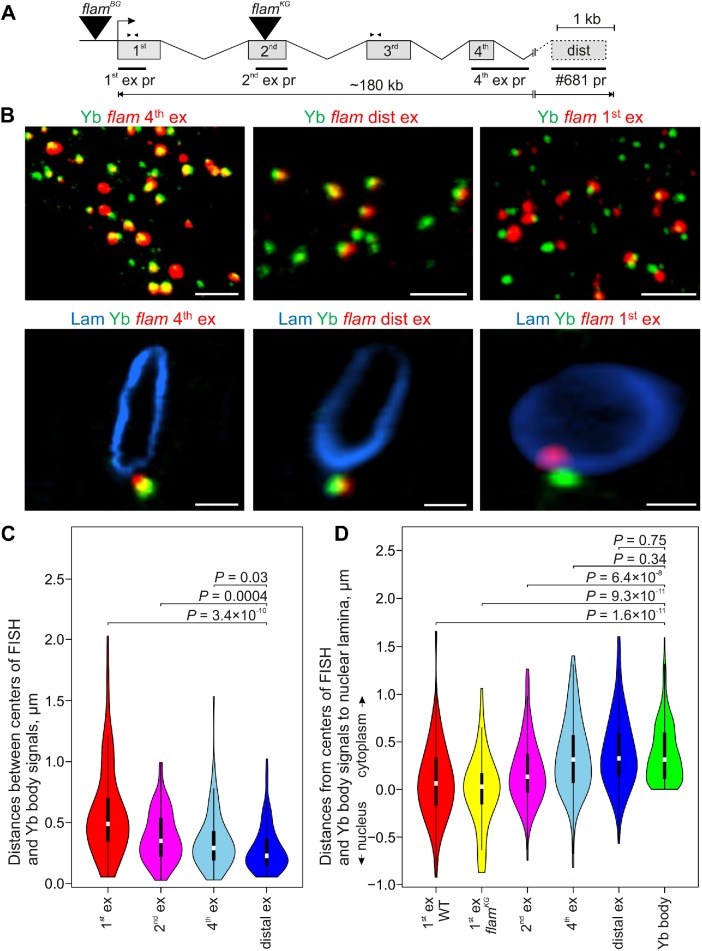
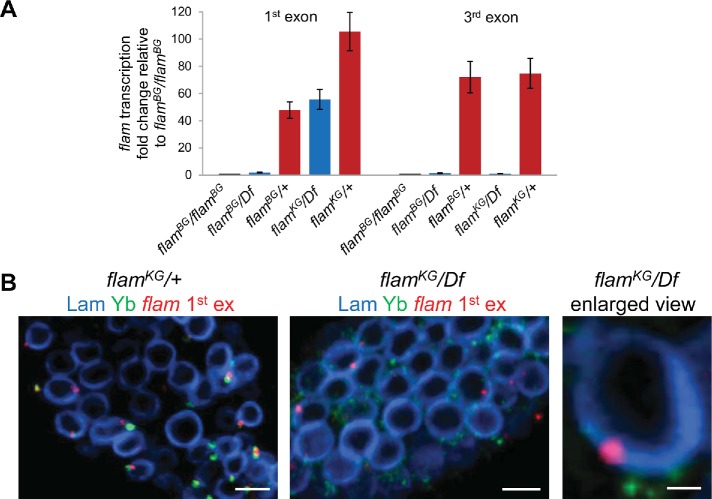
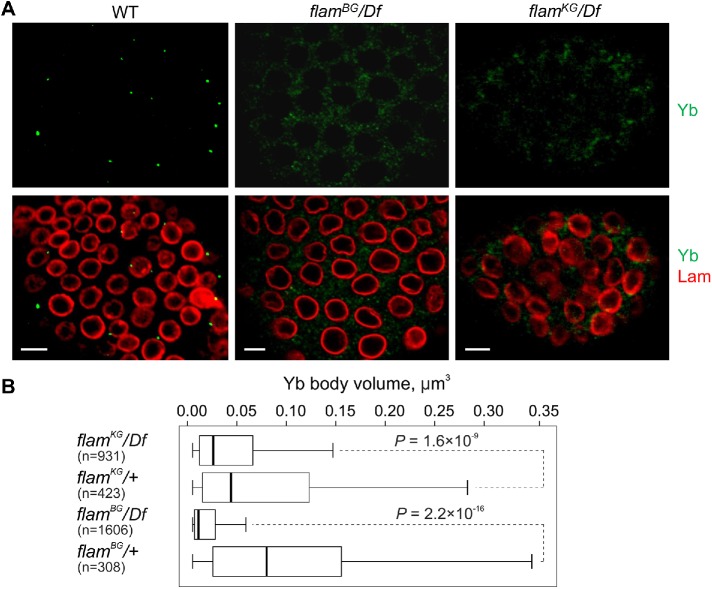
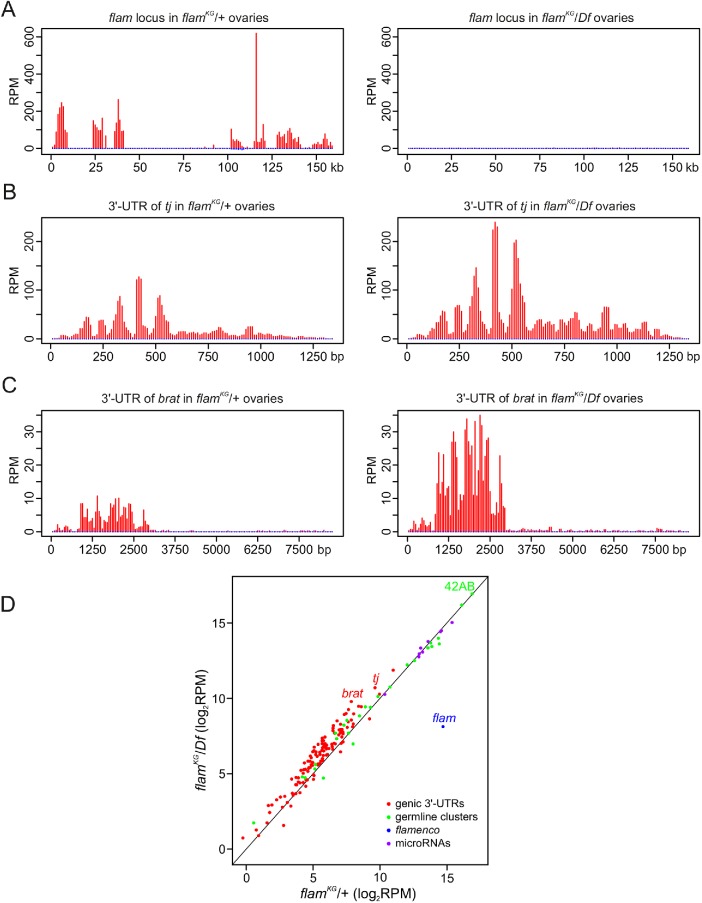
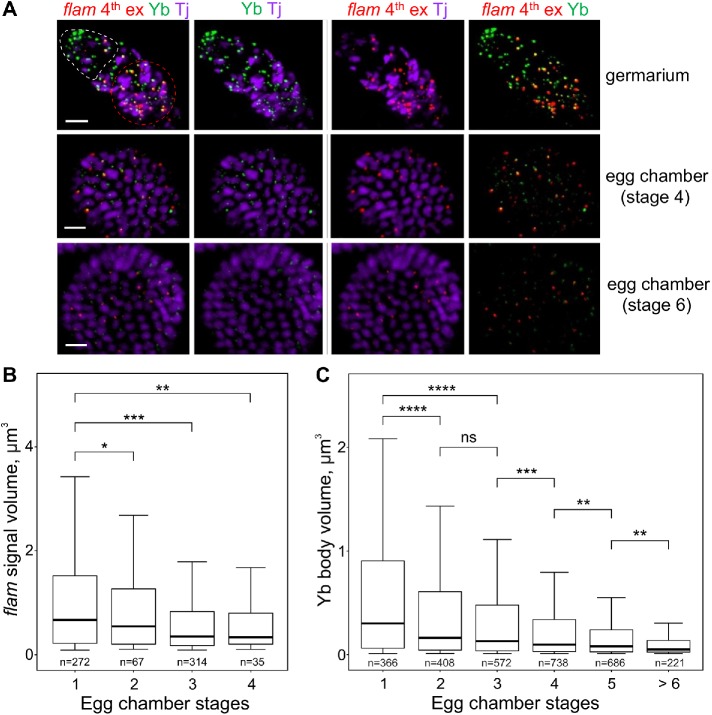
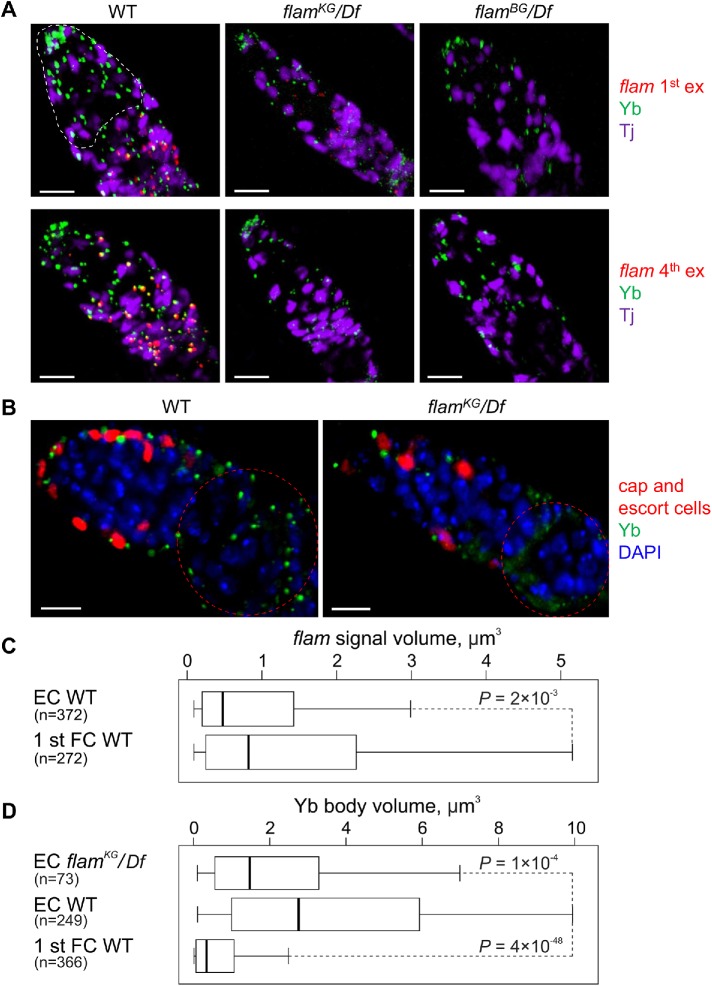
In Drosophila ovarian somatic cells, PIWI-interacting small RNAs (piRNAs) against transposable elements are mainly produced from the ∼180-kb flamenco (flam) locus. flam transcripts are gathered into foci, located close to the nuclear envelope, and processed into piRNAs in the cytoplasmic Yb bodies. The mechanism of Yb body formation remains unknown. Using RNA fluorescence in situ hybridization, we found that in the follicle cells of ovaries the 5'-ends of flam transcripts are usually located in close proximity to the nuclear envelope and outside of Yb bodies, whereas their extended downstream regions mostly overlap with Yb bodies. In flamKG mutant ovaries, flam transcripts containing the first and, partially, second exons but lacking downstream regions are gathered into foci at the nuclear envelope, but Yb bodies are not assembled. Strikingly, piRNAs from the protein-coding gene transcripts accumulate at higher levels in flamKG ovaries indicating that piRNA biogenesis may occur without Yb bodies. We propose that normally in follicle cells, flam downstream transcript regions function not only as a substrate for generation of piRNAs but also as a scaffold for Yb body assembly, which competitively decreases piRNA production from the protein-coding gene transcripts. By contrast, in ovarian somatic cap and escort cells Yb body assembly does not require flam transcription.
Figures
References
-
- Brennecke J, Aravin AA, Stark A, Dus M, Kellis M, Sachidanandam R, Hannon GJ. (2007). Discrete small RNA-generating loci as master regulators of transposon activity in Drosophila . Cell , 1089–1103. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
