

USP49 negatively regulates cellular antiviral responses via deconjugating K63-linked ubiquitination of MITA
- PMID: 30943264
- PMCID: PMC6464240
- DOI: 10.1371/journal.ppat.1007680
USP49 negatively regulates cellular antiviral responses via deconjugating K63-linked ubiquitination of MITA
Abstract
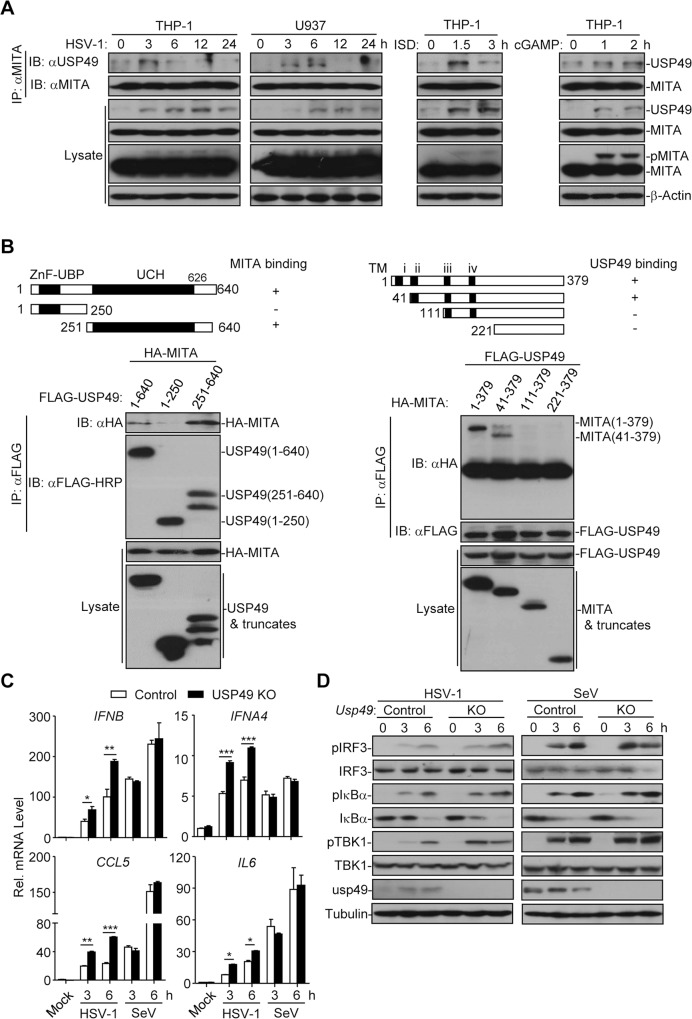
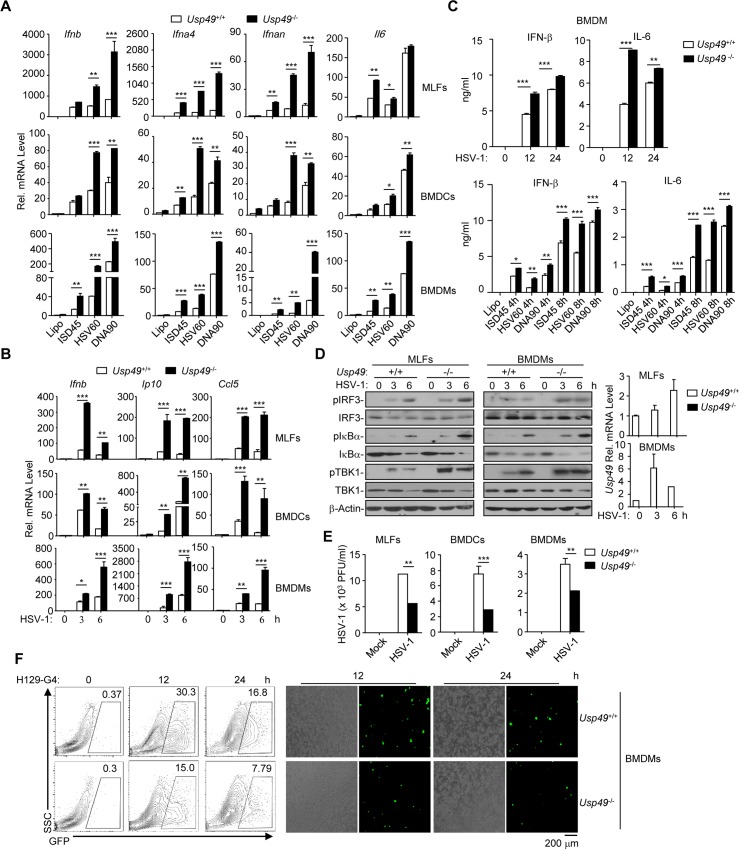
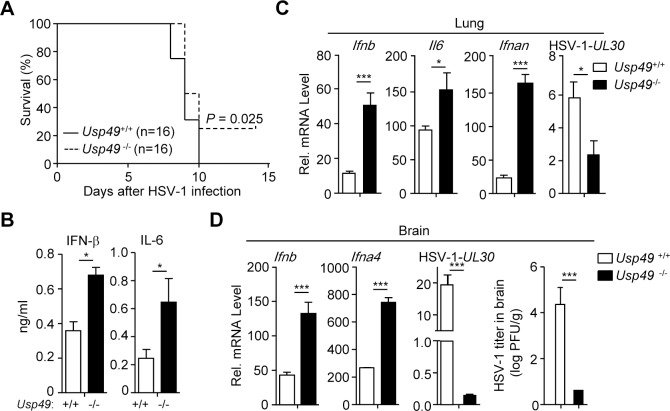
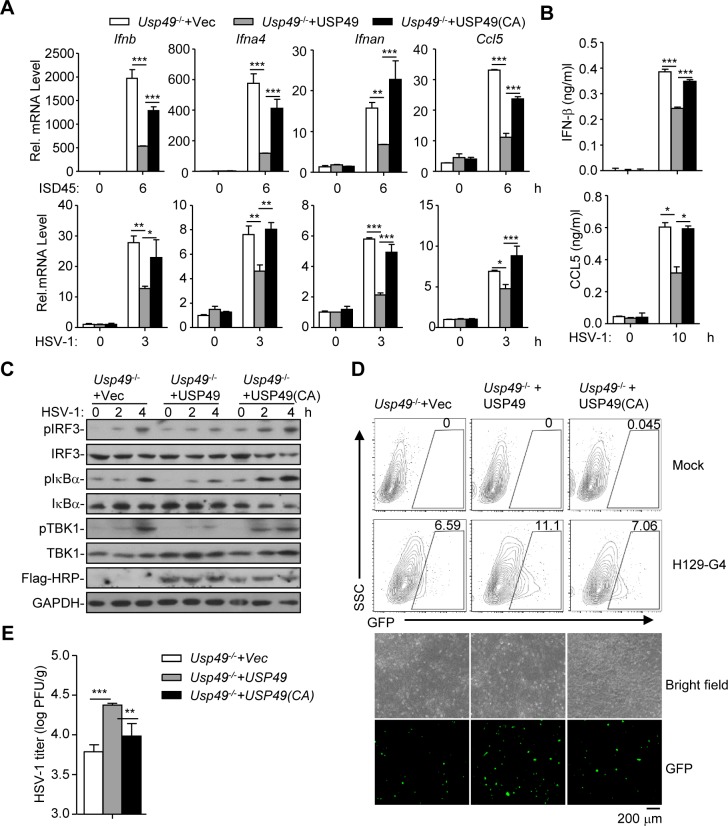
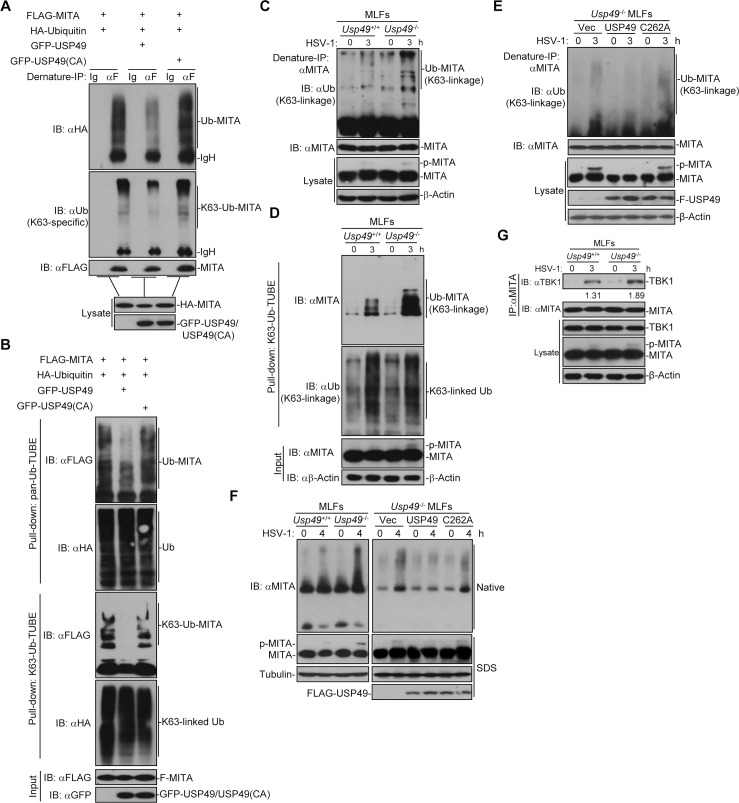
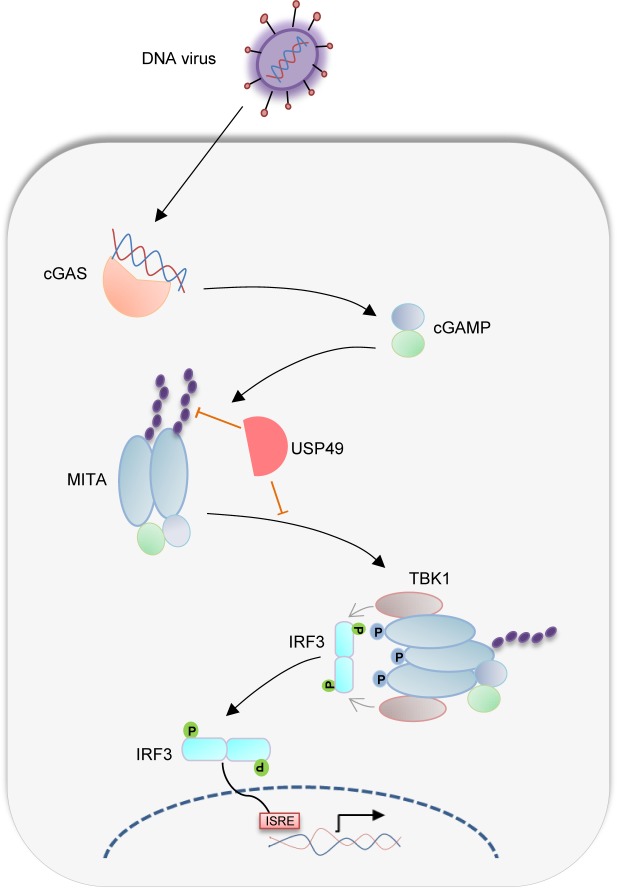
Mediator of IRF3 activation (MITA, also known as STING and ERIS) is an essential adaptor protein for cytoplasmic DNA-triggered signaling and involved in innate immune responses, autoimmunity and tumorigenesis. The activity of MITA is critically regulated by ubiquitination and deubiquitination. Here, we report that USP49 interacts with and deubiquitinates MITA after HSV-1 infection, thereby turning down cellular antiviral responses. Knockdown or knockout of USP49 potentiated HSV-1-, cytoplasmic DNA- or cGAMP-induced production of type I interferons (IFNs) and proinflammatory cytokines and impairs HSV-1 replication. Consistently, Usp49-/- mice exhibit resistance to lethal HSV-1 infection and attenuated HSV-1 replication compared to Usp49+/+ mice. Mechanistically, USP49 removes K63-linked ubiquitin chains from MITA after HSV-1 infection which inhibits the aggregation of MITA and the subsequent recruitment of TBK1 to the signaling complex. These findings suggest a critical role of USP49 in terminating innate antiviral responses and provide insights into the complex regulatory mechanisms of MITA activation.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
