

Watch What You (Self-) Eat: Autophagic Mechanisms that Modulate Metabolism
- PMID: 30943392
- PMCID: PMC6450419
- DOI: 10.1016/j.cmet.2019.03.003
Watch What You (Self-) Eat: Autophagic Mechanisms that Modulate Metabolism
Abstract
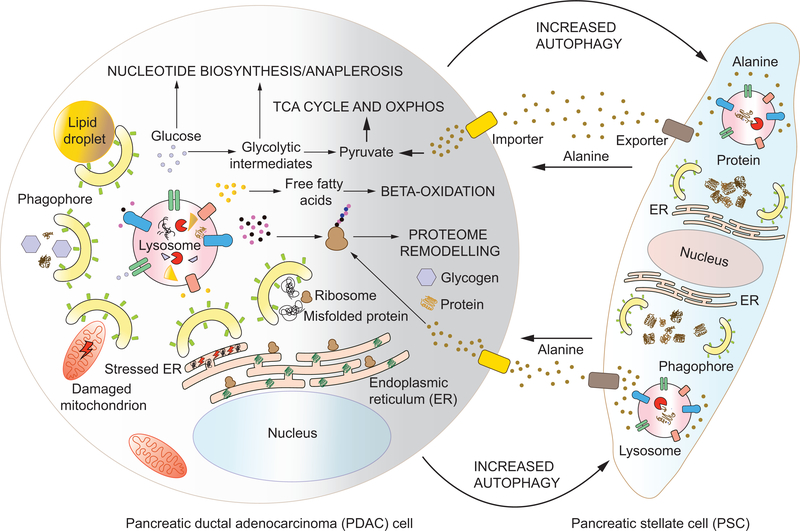
Autophagy is an evolutionarily conserved lysosome- or vacuole-dependent catabolic pathway in eukaryotes. Autophagy functions basally for cellular quality control and is induced to act as an alternative source of basic metabolites during nutrient deprivation. These functions of autophagy are intimately connected to the regulation of metabolism, and the metabolic status of the cell in turn controls the nature and extent of autophagic induction. Here, we highlight the co-regulation of autophagy and metabolism with a special focus on selective autophagy that, along with bulk autophagy, plays a central role in regulating and rewiring metabolic circuits. We outline the metabolic signals that activate these pathways, the mechanisms involved, and the downstream effects and implications while recognizing yet unanswered questions. We also discuss the role of autophagy in the development and maintenance of adipose tissue, an emerging player in systemic metabolic homeostasis, and describe what is currently known about the complex relationship between autophagy and cancer.
Keywords: AMPK; autophagy; ferritinophagy; homeostasis; lysosome; mTOR; macroautophagy; mitophagy; physiology; protein turnover; proteostasis; stress.
Copyright © 2019 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interest
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
