

Innate lymphoid cells support regulatory T cells in the intestine through interleukin-2
- PMID: 30944470
- PMCID: PMC6481643
- DOI: 10.1038/s41586-019-1082-x
Innate lymphoid cells support regulatory T cells in the intestine through interleukin-2
Abstract
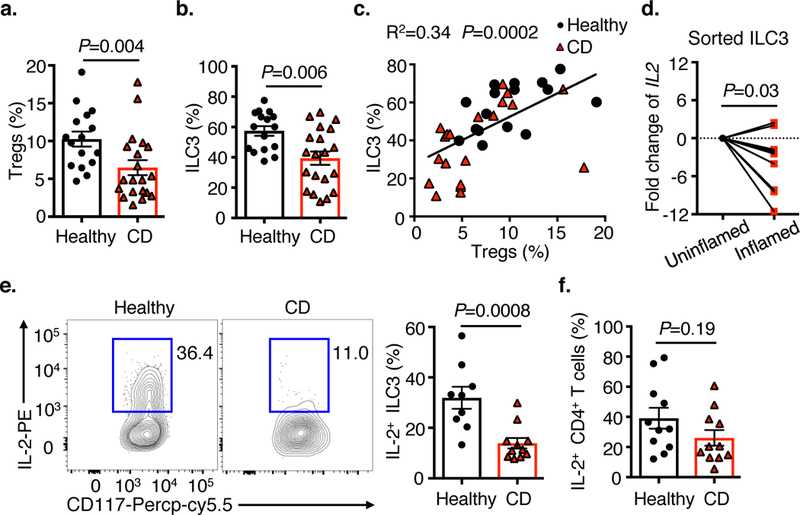
Interleukin (IL)-2 is a pleiotropic cytokine that is necessary to prevent chronic inflammation in the gastrointestinal tract1-4. The protective effects of IL-2 involve the generation, maintenance and function of regulatory T (Treg) cells4-8, and the use of low doses of IL-2 has emerged as a potential therapeutic strategy for patients with inflammatory bowel disease9. However, the cellular and molecular pathways that control the production of IL-2 in the context of intestinal health are undefined. Here we show, in a mouse model, that IL-2 is acutely required to maintain Treg cells and immunological homeostasis throughout the gastrointestinal tract. Notably, lineage-specific deletion of IL-2 in T cells did not reduce Treg cells in the small intestine. Unbiased analyses revealed that, in the small intestine, group-3 innate lymphoid cells (ILC3s) are the dominant cellular source of IL-2, which is induced selectively by IL-1β. Macrophages in the small intestine produce IL-1β, and activation of this pathway involves MYD88- and NOD2-dependent sensing of the microbiota. Our loss-of-function studies show that ILC3-derived IL-2 is essential for maintaining Treg cells, immunological homeostasis and oral tolerance to dietary antigens in the small intestine. Furthermore, production of IL-2 by ILC3s was significantly reduced in the small intestine of patients with Crohn's disease, and this correlated with lower frequencies of Treg cells. Our results reveal a previously unappreciated pathway in which a microbiota- and IL-1β-dependent axis promotes the production of IL-2 by ILC3s to orchestrate immune regulation in the intestine.
Conflict of interest statement
The authors declare no competing interests.
Figures













Comment in
-
ILC3s take control in small intestine.Nat Rev Immunol. 2019 Jun;19(6):353. doi: 10.1038/s41577-019-0166-z. Nat Rev Immunol. 2019. PMID: 30979958 No abstract available.
References
-
- Sadlack B et al. Ulcerative colitis-like disease in mice with a disrupted interleukin-2 gene. Cell 75, 253–261 (1993). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
