

GPCR-independent activation of G proteins promotes apical cell constriction in vivo
- PMID: 30948426
- PMCID: PMC6504902
- DOI: 10.1083/jcb.201811174
GPCR-independent activation of G proteins promotes apical cell constriction in vivo
Abstract
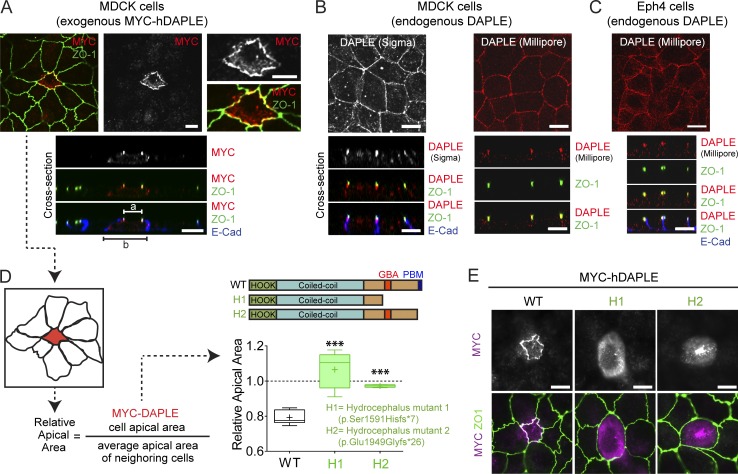
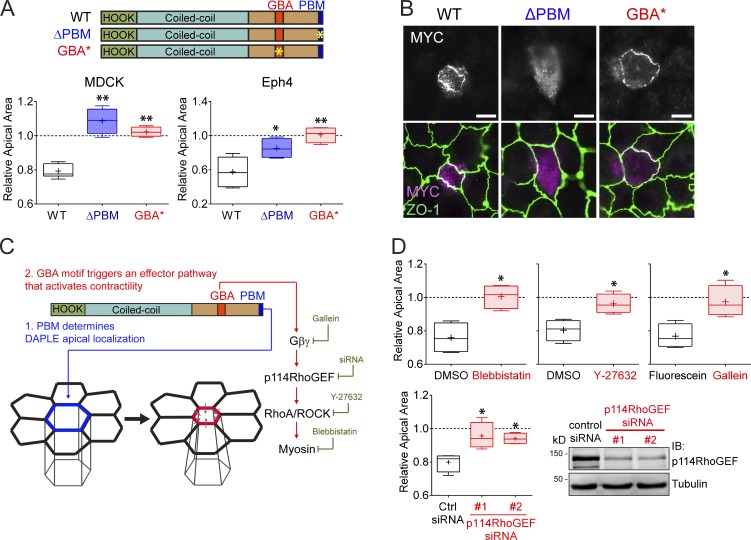
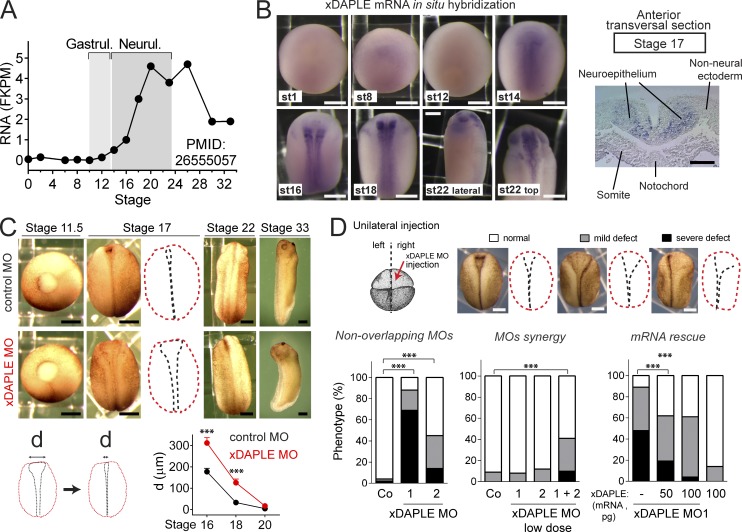
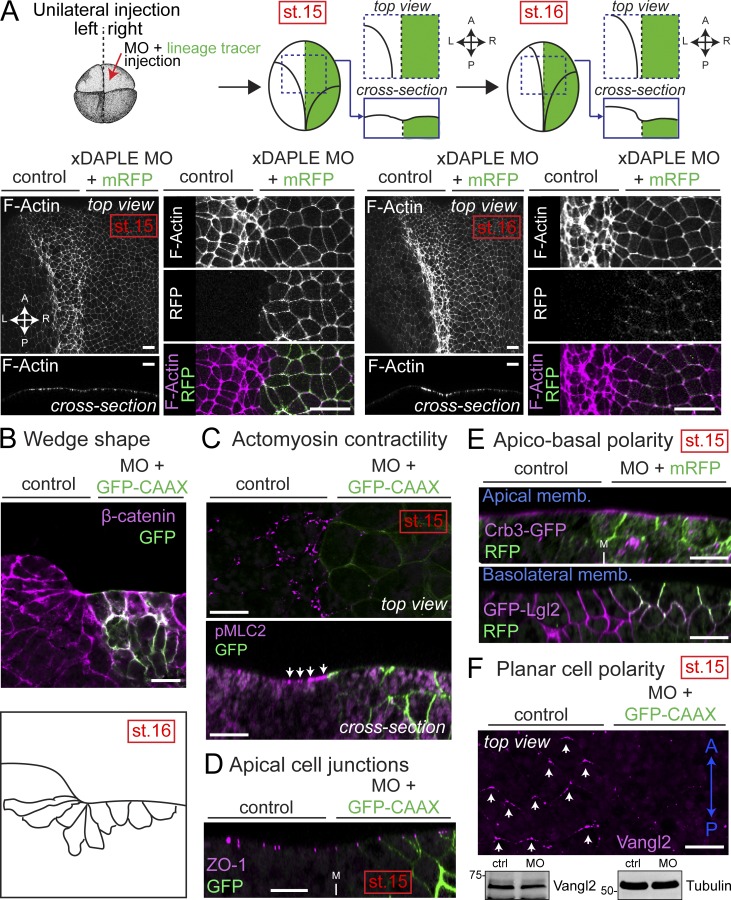
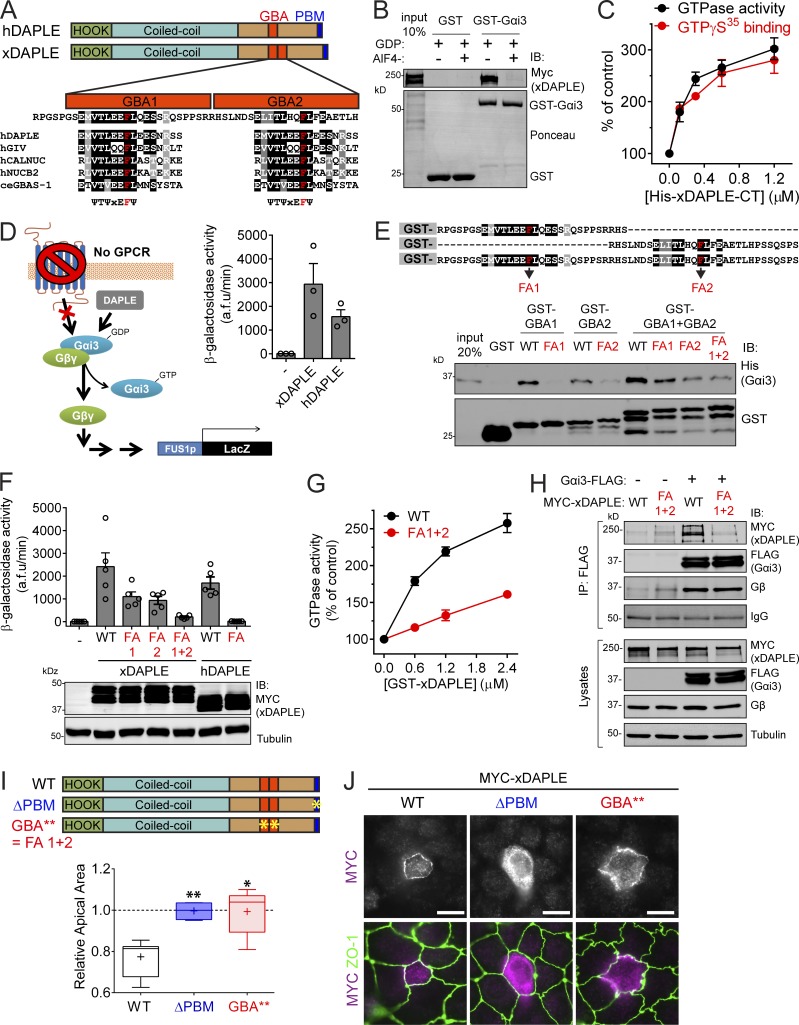
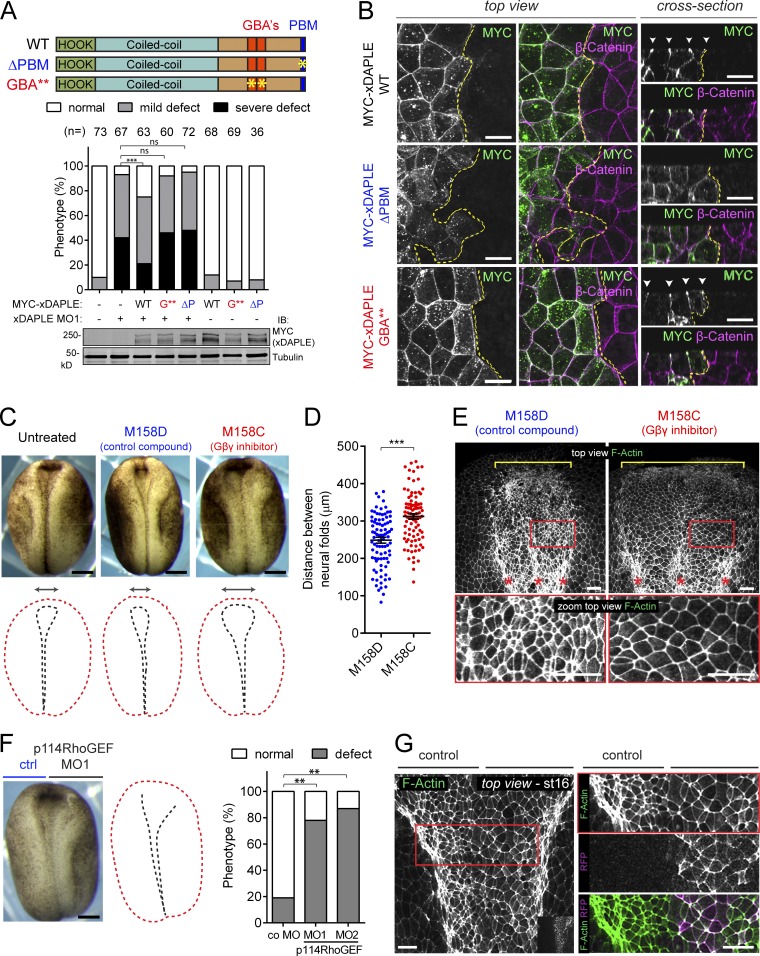
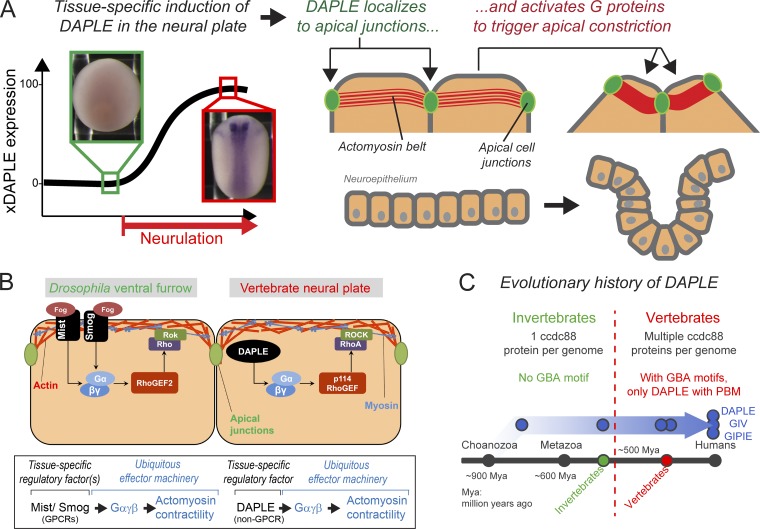
Heterotrimeric G proteins are signaling switches that control organismal morphogenesis across metazoans. In invertebrates, specific GPCRs instruct G proteins to promote collective apical cell constriction in the context of epithelial tissue morphogenesis. In contrast, tissue-specific factors that instruct G proteins during analogous processes in vertebrates are largely unknown. Here, we show that DAPLE, a non-GPCR protein linked to human neurodevelopmental disorders, is expressed specifically in the neural plate of Xenopus laevis embryos to trigger a G protein signaling pathway that promotes apical cell constriction during neurulation. DAPLE localizes to apical cell-cell junctions in the neuroepithelium, where it activates G protein signaling to drive actomyosin-dependent apical constriction and subsequent bending of the neural plate. This function is mediated by a Gα-binding-and-activating (GBA) motif that was acquired by DAPLE in vertebrates during evolution. These findings reveal that regulation of tissue remodeling during vertebrate development can be driven by an unconventional mechanism of heterotrimeric G protein activation that operates in lieu of GPCRs.
© 2019 Marivin et al.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
