

Oncogenic PIK3CA promotes cellular stemness in an allele dose-dependent manner
- PMID: 30948643
- PMCID: PMC6486754
- DOI: 10.1073/pnas.1821093116
Oncogenic PIK3CA promotes cellular stemness in an allele dose-dependent manner
Abstract
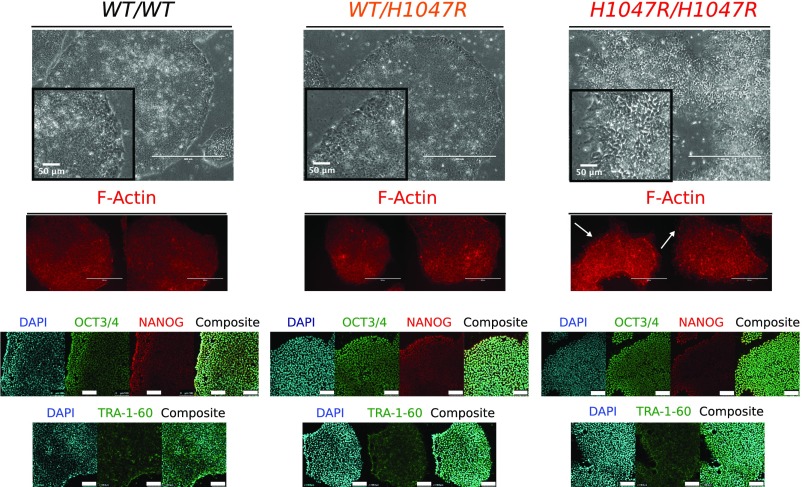
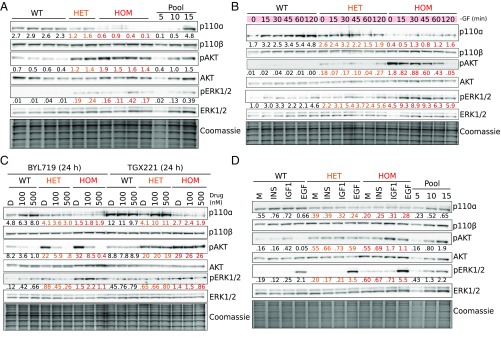
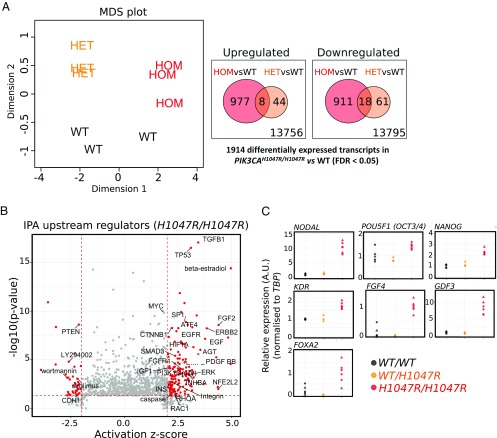
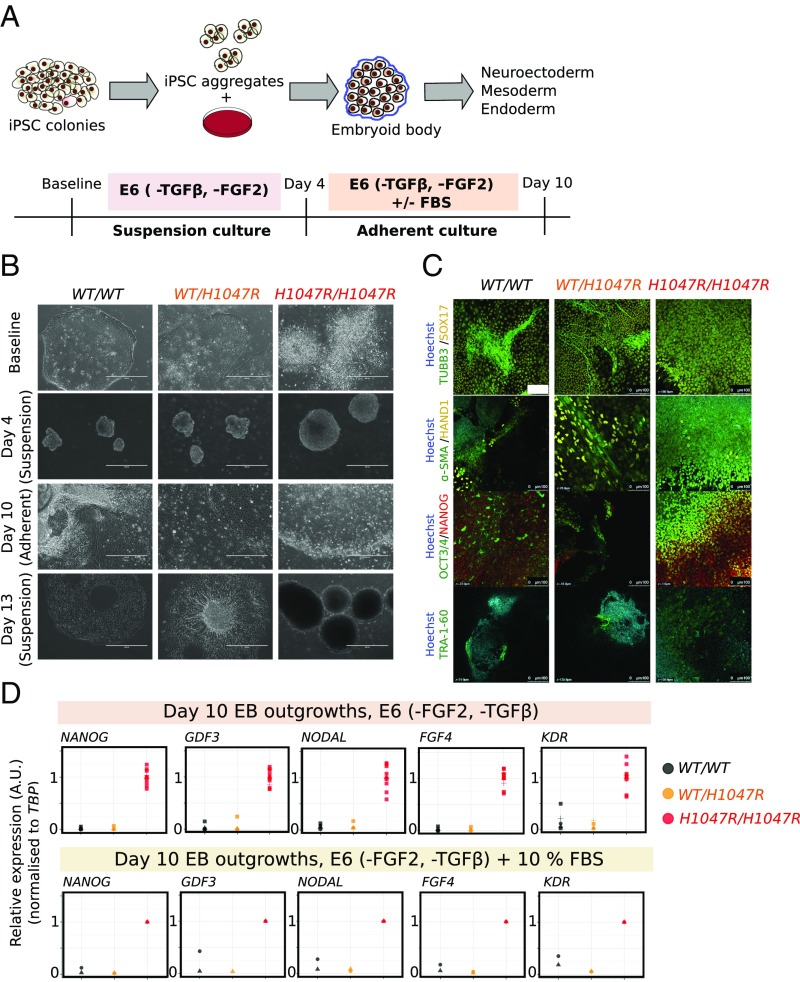
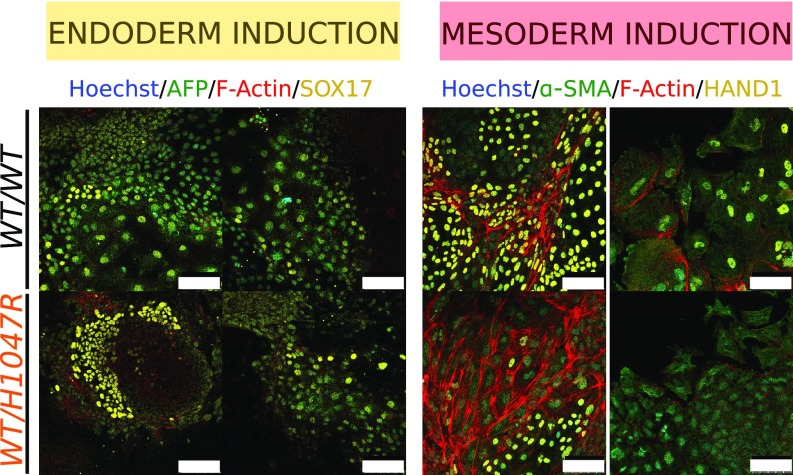
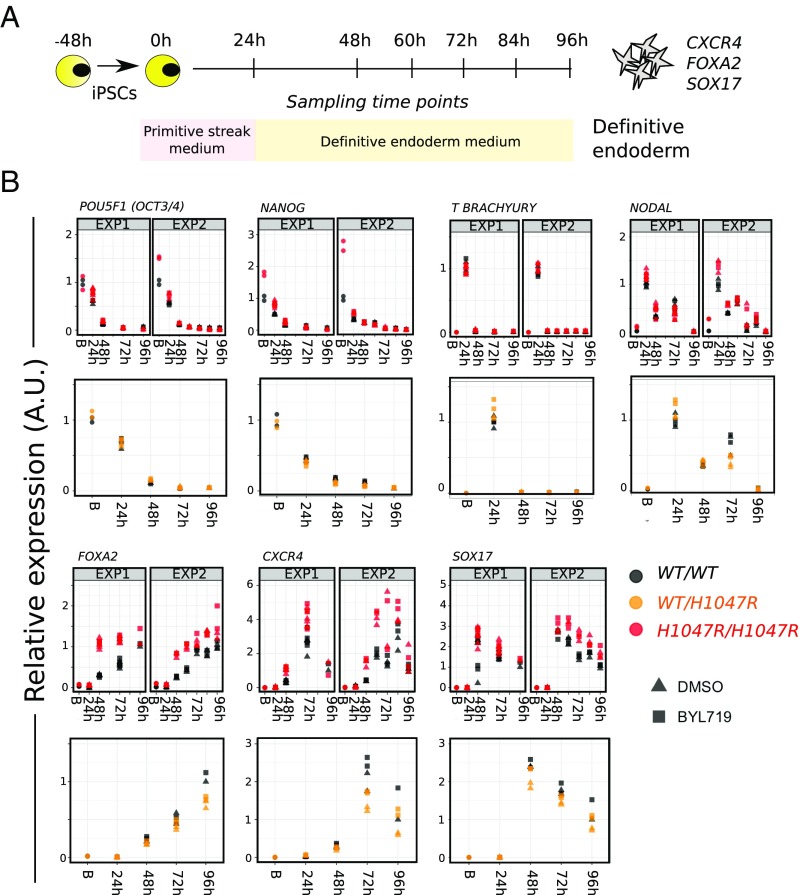
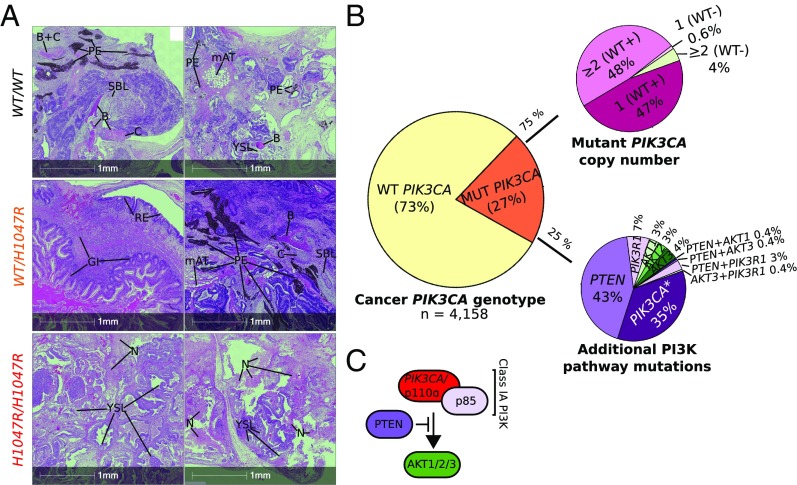
The PIK3CA gene, which encodes the p110α catalytic subunit of PI3 kinase (PI3K), is mutationally activated in cancer and in overgrowth disorders known as PIK3CA-related overgrowth spectrum (PROS). To determine the consequences of genetic PIK3CA activation in a developmental context of relevance to both PROS and cancer, we engineered isogenic human induced pluripotent stem cells (iPSCs) with heterozygous or homozygous knockin of PIK3CAH1047R While heterozygous iPSCs remained largely similar to wild-type cells, homozygosity for PIK3CAH1047R caused widespread, cancer-like transcriptional remodeling, partial loss of epithelial morphology, up-regulation of stemness markers, and impaired differentiation to all three germ layers in vitro and in vivo. Genetic analysis of PIK3CA-associated cancers revealed that 64% had multiple oncogenic PIK3CA copies (39%) or additional PI3K signaling pathway-activating "hits" (25%). This contrasts with the prevailing view that PIK3CA mutations occur heterozygously in cancer. Our findings suggest that a PI3K activity threshold determines pathological consequences of oncogenic PIK3CA activation and provide insight into the specific role of this pathway in human pluripotent stem cells.
Keywords: PI3K; PROS; cancer; genetics; pluripotent stem cells.
Copyright © 2019 the Author(s). Published by PNAS.
Conflict of interest statement
Conflict of interest statement: B.V. is a consultant for Venthera (Palo Alto, CA), iOnctura (Geneva, Switzerland), and Karus Therapeutics (Oxford, United Kingdom). N.M. has received consultancy fees from Achilles Therapeutics.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
