

Establishing and regulating the composition of cilia for signal transduction
- PMID: 30948801
- PMCID: PMC6738346
- DOI: 10.1038/s41580-019-0116-4
Establishing and regulating the composition of cilia for signal transduction
Abstract
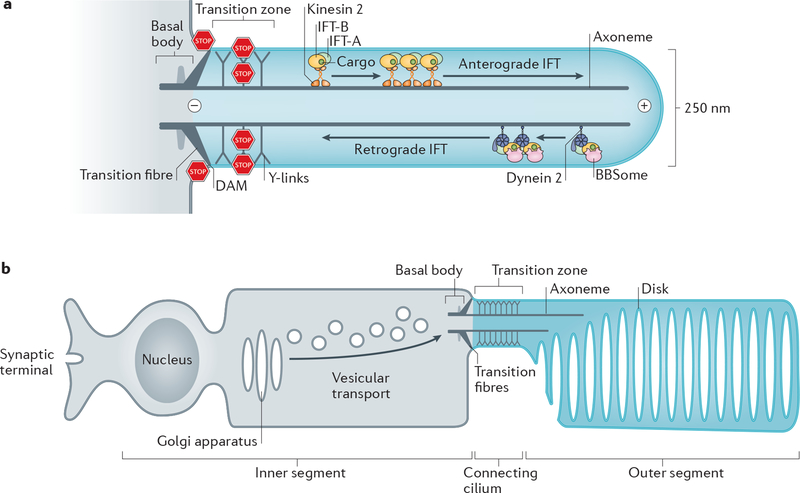
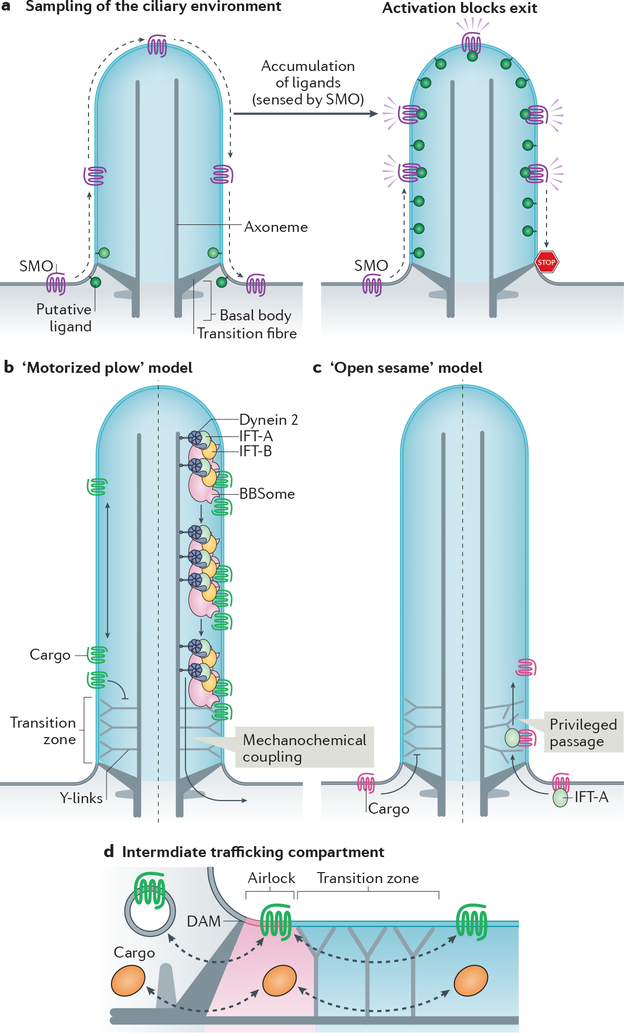
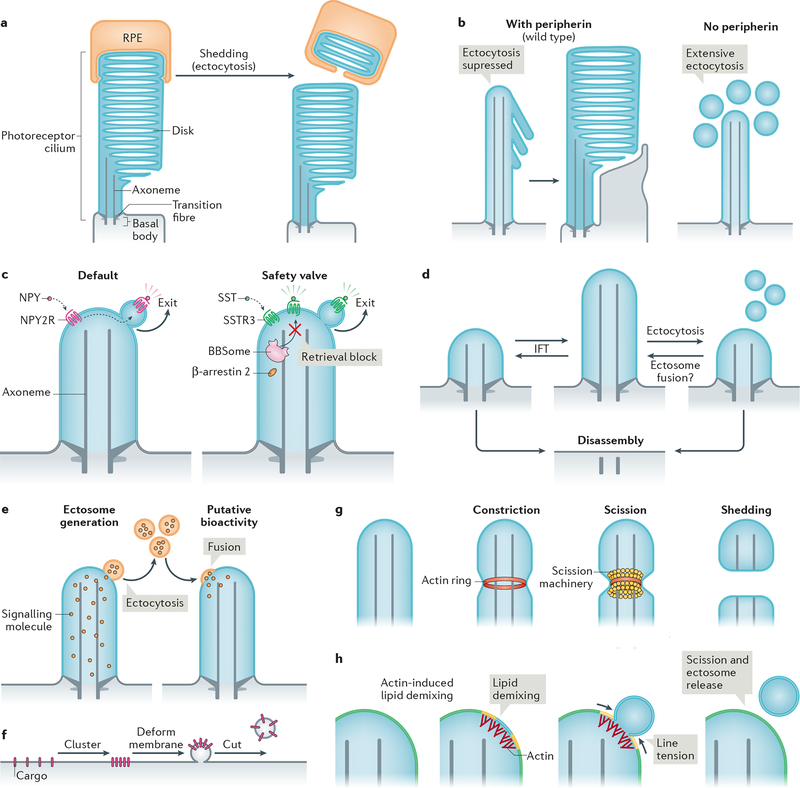
The primary cilium is a hair-like surface-exposed organelle of the eukaryotic cell that decodes a variety of signals - such as odorants, light and Hedgehog morphogens - by altering the local concentrations and activities of signalling proteins. Signalling within the cilium is conveyed through a diverse array of second messengers, including conventional signalling molecules (such as cAMP) and some unusual intermediates (such as sterols). Diffusion barriers at the ciliary base establish the unique composition of this signalling compartment, and cilia adapt their proteome to signalling demands through regulated protein trafficking. Much progress has been made on the molecular understanding of regulated ciliary trafficking, which encompasses not only exchanges between the cilium and the rest of the cell but also the shedding of signalling factors into extracellular vesicles.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures

References
-
- Bloom G Studies on the olfactory epithelium of the frog and the toad with the aid of light and electron microscopy. Zeitschrift für Zellforschung und Mikroskopische Anatomie 41, 89–100 (1954). - PubMed
-
- Huangfu D et al. Hedgehog signalling in the mouse requires intraflagellar transport proteins. Nature 426, 83–7 (2003). - PubMed
-
- Gerdes JM et al. Disruption of the basal body compromises proteasomal function and perturbs intracellular Wnt response. Nature Genetics 39, 1350–1360 (2007). - PubMed
