

Reassessment of Exosome Composition
- PMID: 30951670
- PMCID: PMC6664447
- DOI: 10.1016/j.cell.2019.02.029
Reassessment of Exosome Composition
Abstract
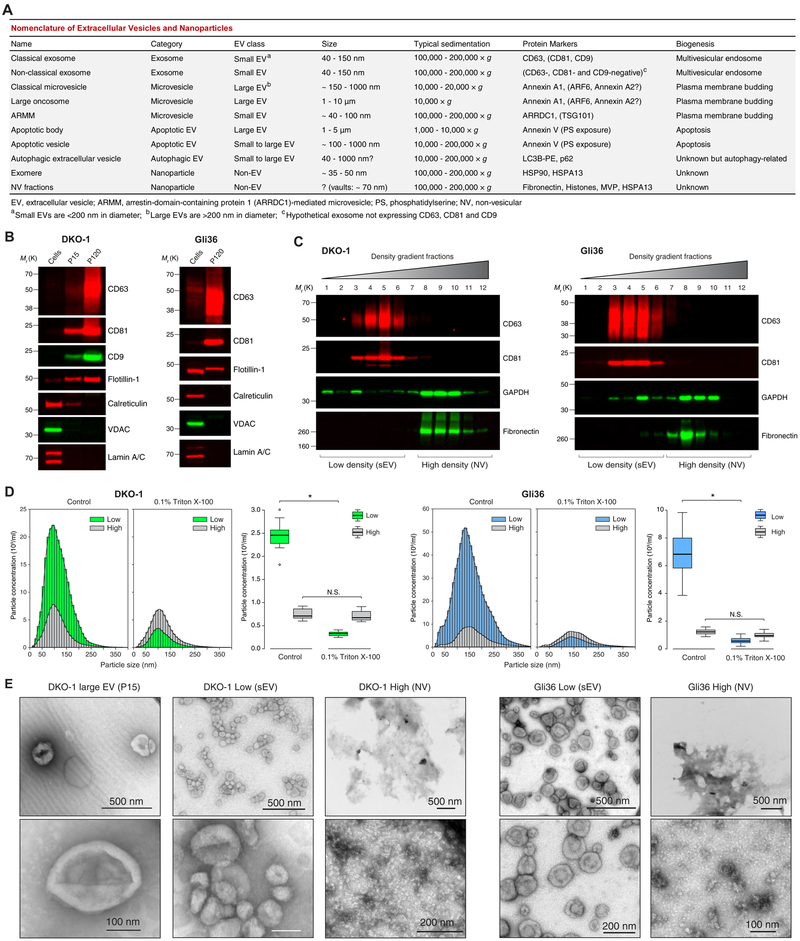
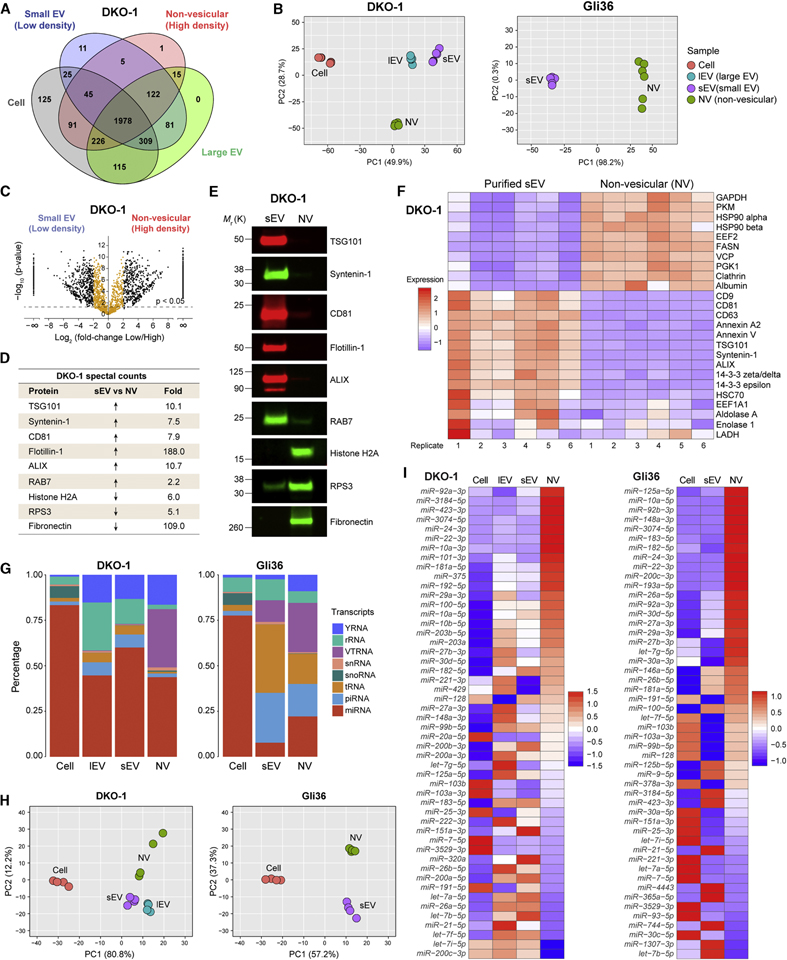
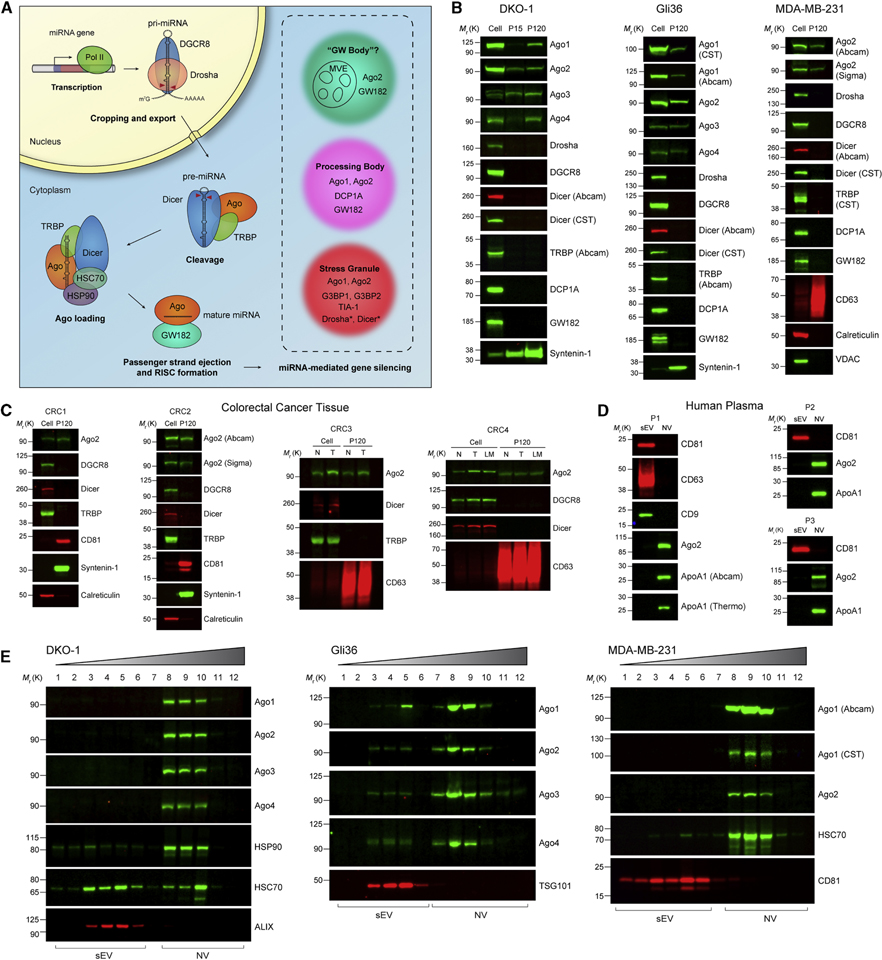
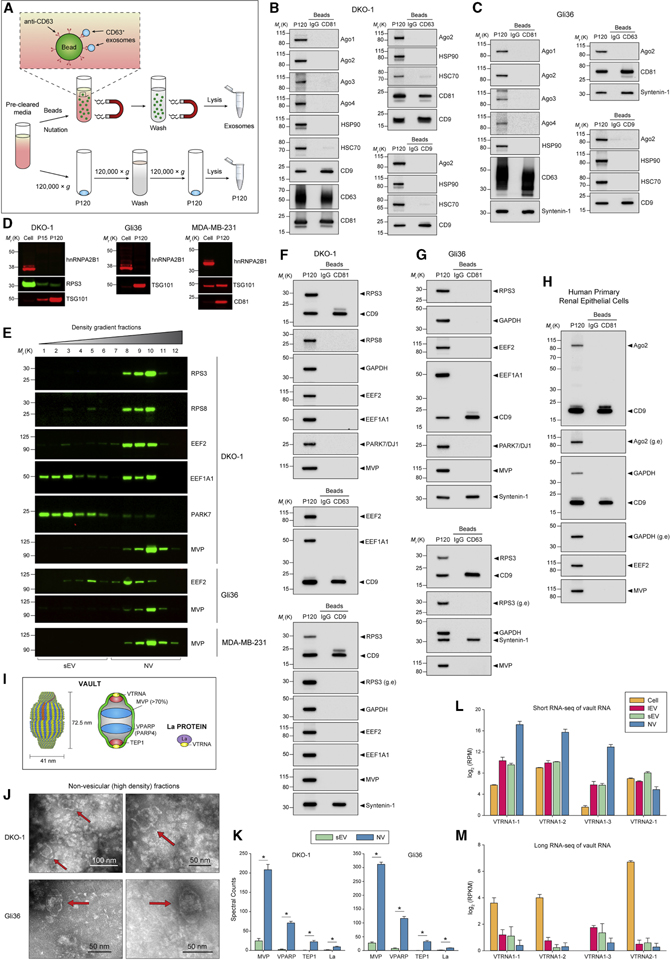
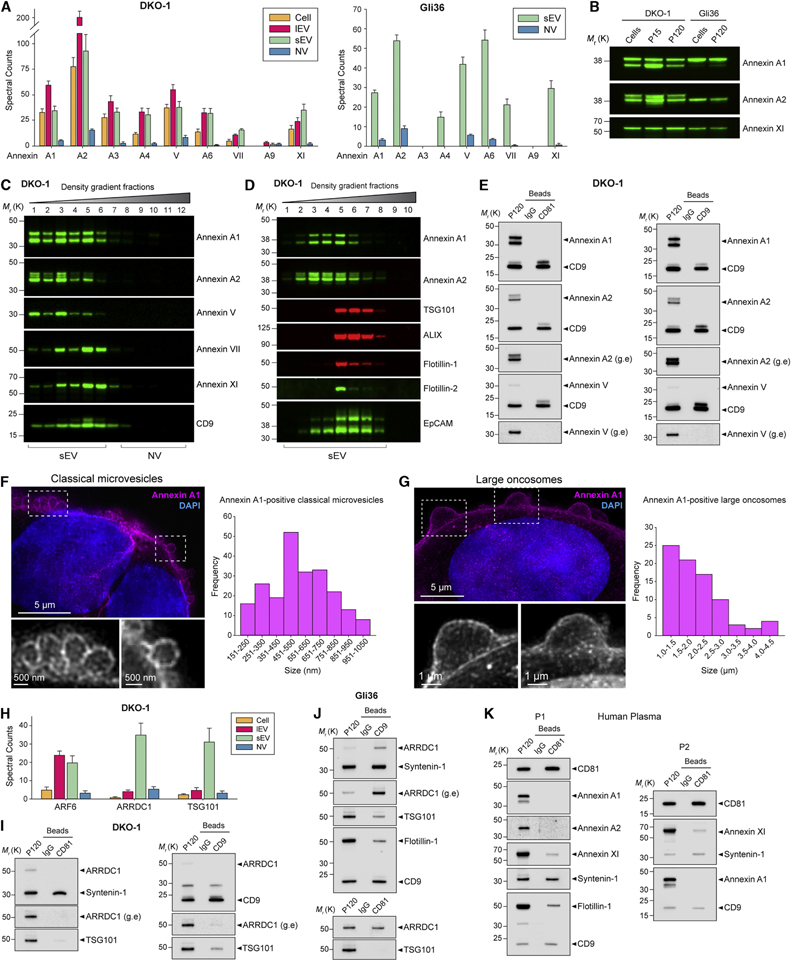
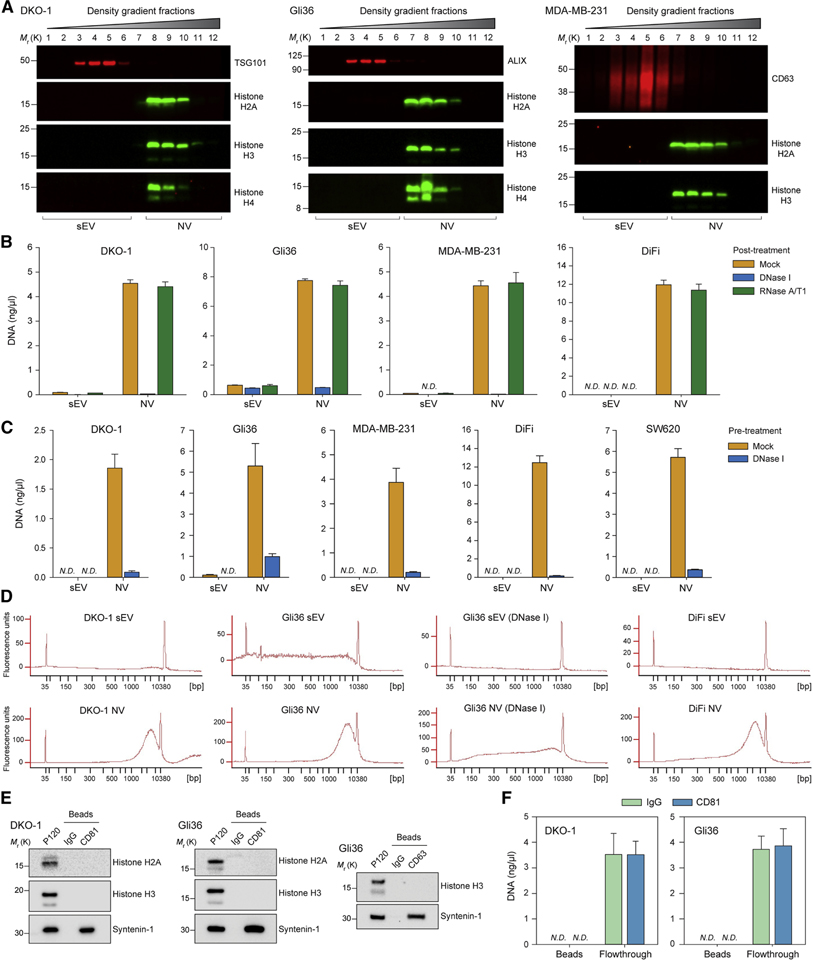
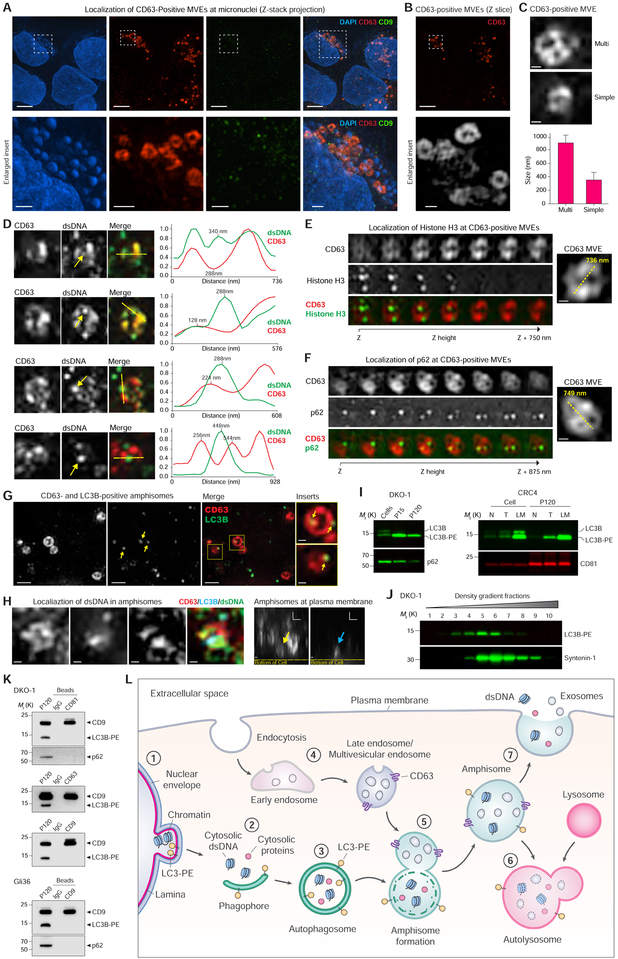
The heterogeneity of small extracellular vesicles and presence of non-vesicular extracellular matter have led to debate about contents and functional properties of exosomes. Here, we employ high-resolution density gradient fractionation and direct immunoaffinity capture to precisely characterize the RNA, DNA, and protein constituents of exosomes and other non-vesicle material. Extracellular RNA, RNA-binding proteins, and other cellular proteins are differentially expressed in exosomes and non-vesicle compartments. Argonaute 1-4, glycolytic enzymes, and cytoskeletal proteins were not detected in exosomes. We identify annexin A1 as a specific marker for microvesicles that are shed directly from the plasma membrane. We further show that small extracellular vesicles are not vehicles of active DNA release. Instead, we propose a new model for active secretion of extracellular DNA through an autophagy- and multivesicular-endosome-dependent but exosome-independent mechanism. This study demonstrates the need for a reassessment of exosome composition and offers a framework for a clearer understanding of extracellular vesicle heterogeneity.
Keywords: Argonaute; amphisomes; annexin; autophagy; exomeres; exosomes; extracellular DNA; extracellular RNA; extracellular vesicles; microvesicles.
Copyright © 2019 Elsevier Inc. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
The authors declare no competing interests.
Figures
Comment in
-
Explicating Exosomes: Reclassifying the Rising Stars of Intercellular Communication.Cell. 2019 Apr 4;177(2):225-227. doi: 10.1016/j.cell.2019.03.020. Cell. 2019. PMID: 30951665
References
-
- Arroyo JD, Chevillet JR, Kroh EM, Ruf IK, Pritchard CC, Gibson DF, Mitchell PS, Bennett CF, Pogosova-Agadjanyan EL, Stirewalt DL, et al. (2011). Argonaute2 complexes carry a population of circulating microRNAs independent of vesicles in human plasma. Proc Natl Acad Sci U S A 108, 5003–5008. - PMC - PubMed
-
- Berg TO, Fengsrud M, Stromhaug PE, Berg T, and Seglen PO (1998). Isolation and characterization of rat liver amphisomes. Evidence for fusion of autophagosomes with both early and late endosomes. J Biol Chem 273, 21883–21892. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
