

Keratinocyte-Mediated Activation of the Cytokine TGF-β Maintains Skin Recirculating Memory CD8+ T Cells
- PMID: 30952606
- PMCID: PMC6531326
- DOI: 10.1016/j.immuni.2019.03.002
Keratinocyte-Mediated Activation of the Cytokine TGF-β Maintains Skin Recirculating Memory CD8+ T Cells
Abstract
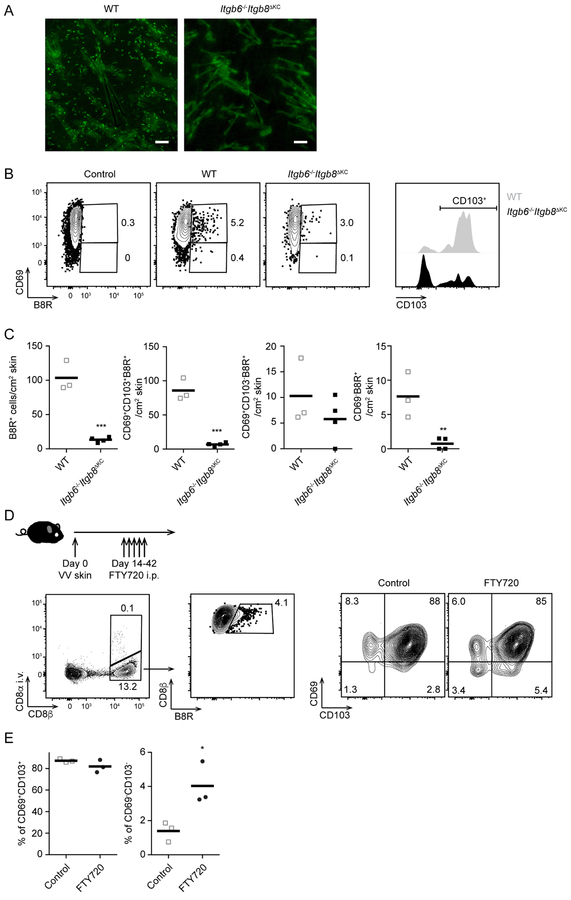
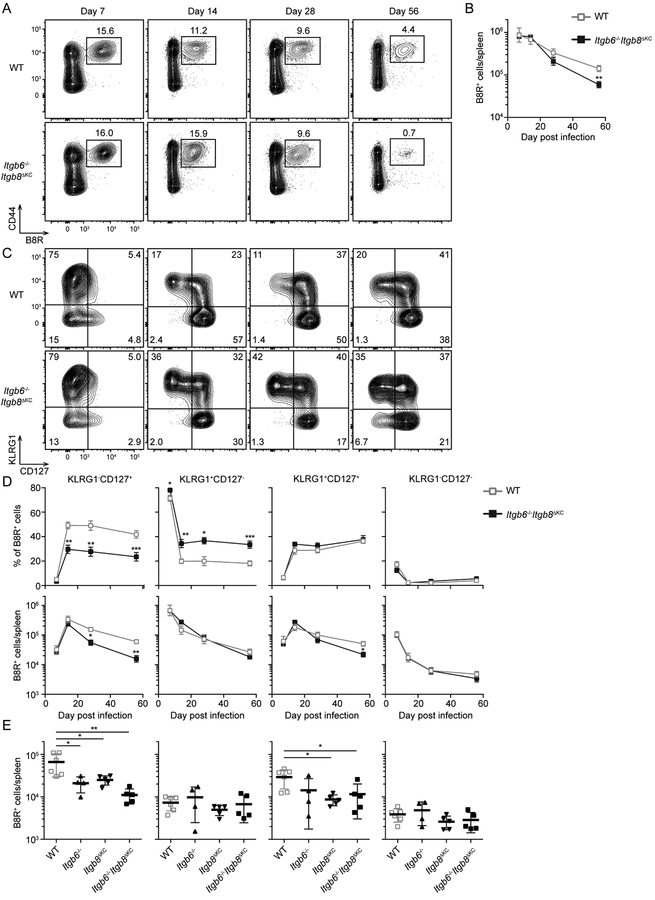
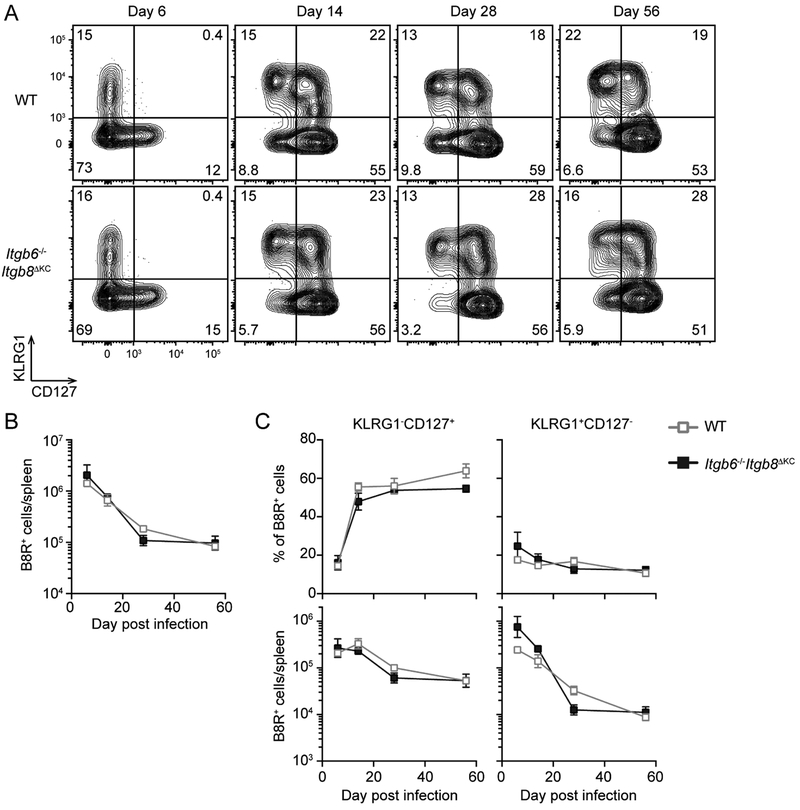
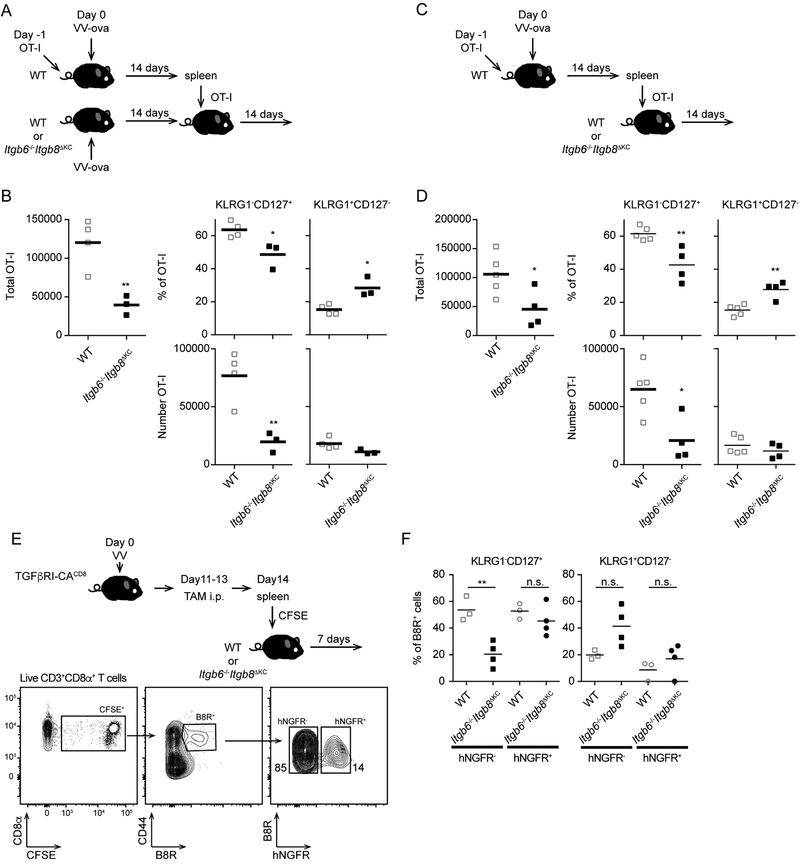
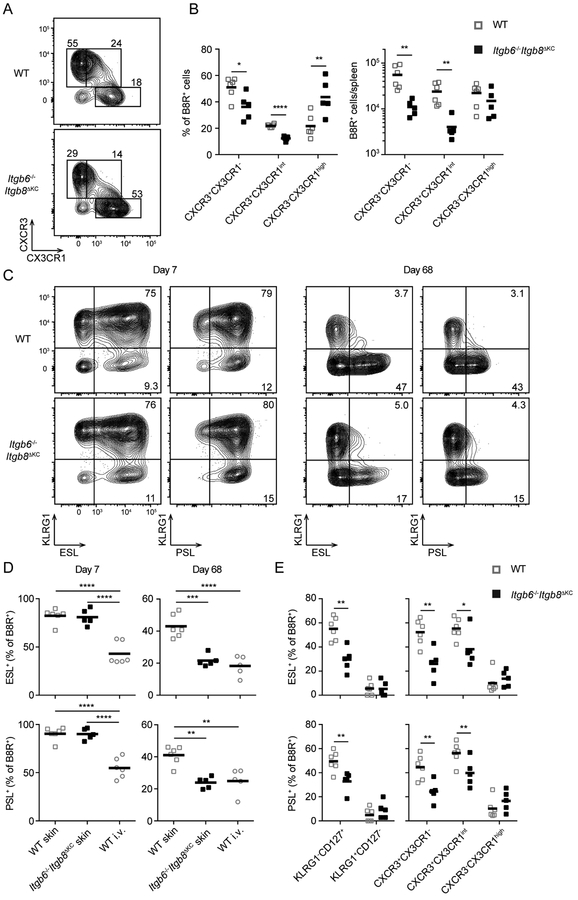
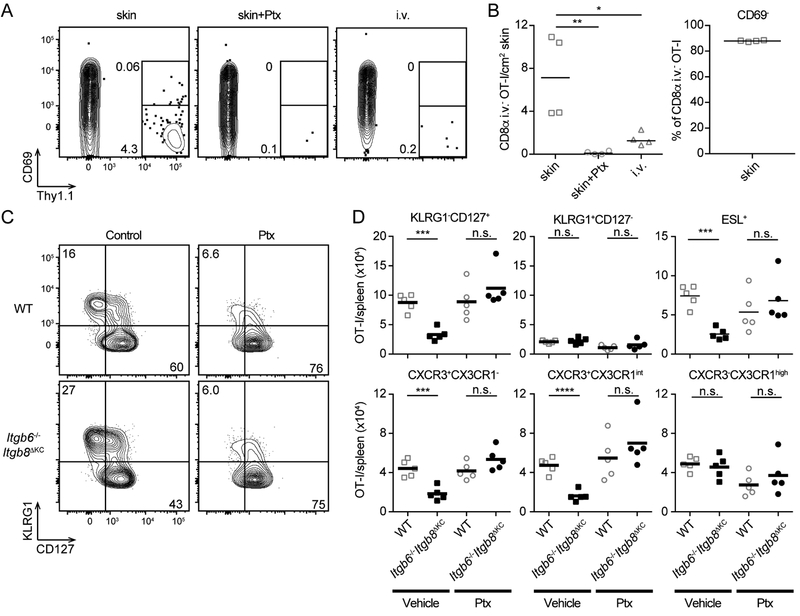
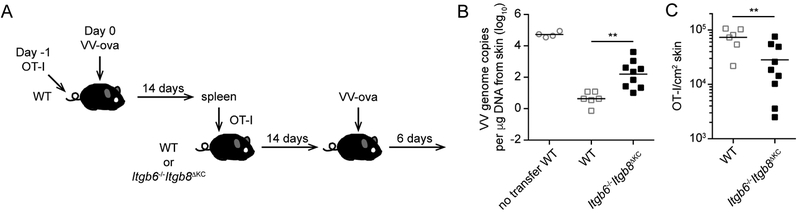
Regulated activation of the cytokine TGF-β by integrins αvβ6 and αvβ8 expressed on keratinocytes is required for residence of epidermal-resident memory T cells, but whether skin-derived signals also affect recirculating memory cells in the skin remains unclear. Here, we show that after resolution of skin vaccinia virus (VV) infection, antigen-specific circulating memory CD8+ T cells migrated into skin. In mice lacking αvβ6 and αvβ8 integrins (Itgb6-/-Itgb8fl/fl-K14-cre), the absence of epidermal-activated TGF-β resulted in a gradual loss of E- or P-selectin-binding central and peripheral memory populations, which were rescued when skin entry was inhibited. Skin recirculating memory cells were required for optimal host defense against skin VV infection. These data demonstrate that skin migration can persist after resolution of local skin infection and that the cytokine environment within this nonlymphoid tissue shapes the differentiation state and persistence of the central and peripheral memory-T-cell pool.
Keywords: CD8(+) T cell memory; keratinocytes; skin; transforming growth factor beta; α(v)β(6); α(v)β(8).
Copyright © 2019 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests
The authors declare no competing interests.
Figures
References
-
- Bartholin L, Cyprian FS, Vincent D, Garcia CN, Martel S, Horvat B, Berthet C, Léon SG, Treilleux I, Rimokh R, Marie JC, 2008. Generation of mice with conditionally activated transforming growth factor beta signaling through the TβRI/ALK5 receptor. genesis 46, 724–731. doi:10.1002/dvg.20425 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
