

Gene signatures of quiescent glioblastoma cells reveal mesenchymal shift and interactions with niche microenvironment
- PMID: 30952620
- PMCID: PMC6491796
- DOI: 10.1016/j.ebiom.2019.03.064
Gene signatures of quiescent glioblastoma cells reveal mesenchymal shift and interactions with niche microenvironment
Abstract
Background: Glioblastoma (GBM), a highly malignant brain tumor, invariably recurs after therapy. Quiescent GBM cells represent a potential source of tumor recurrence, but little is known about their molecular underpinnings.
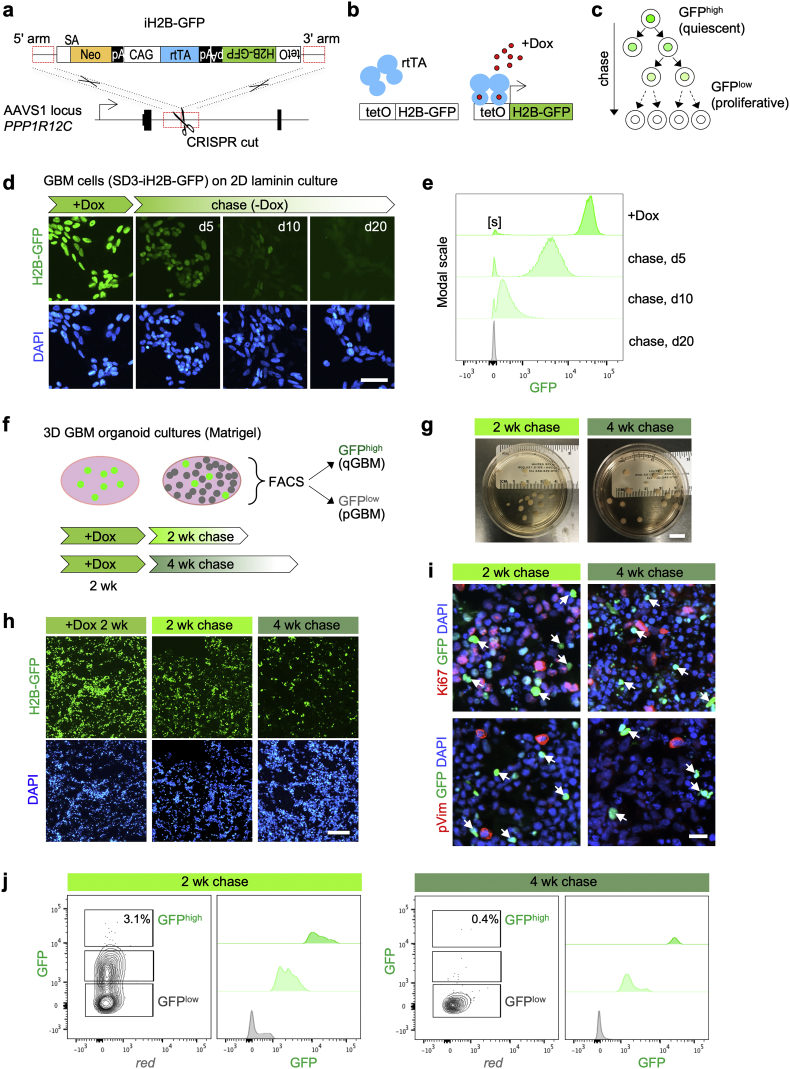
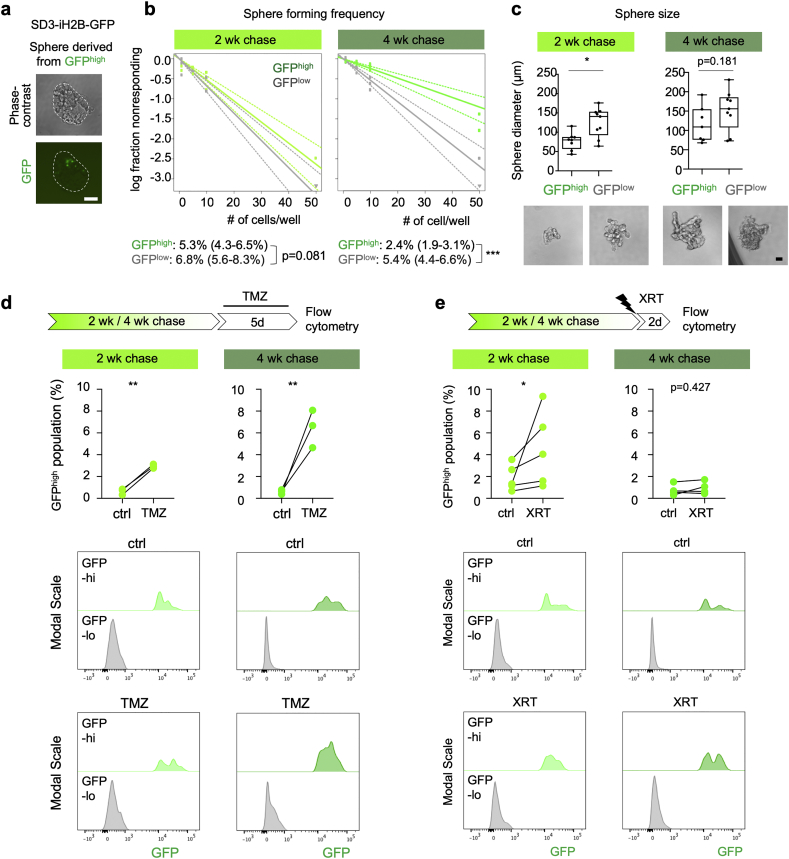
Methods: Patient-derived GBM cells were engineered by CRISPR/Cas9-assisted knock-in of an inducible histone2B-GFP (iH2B-GFP) reporter to track cell division history. We utilized an in vitro 3D GBM organoid approach to isolate live quiescent GBM (qGBM) cells and their proliferative counterparts (pGBM) to compare stem cell properties and therapy resistance. Gene expression programs of qGBM and pGBM cells were analyzed by RNA-Seq and NanoString platforms.
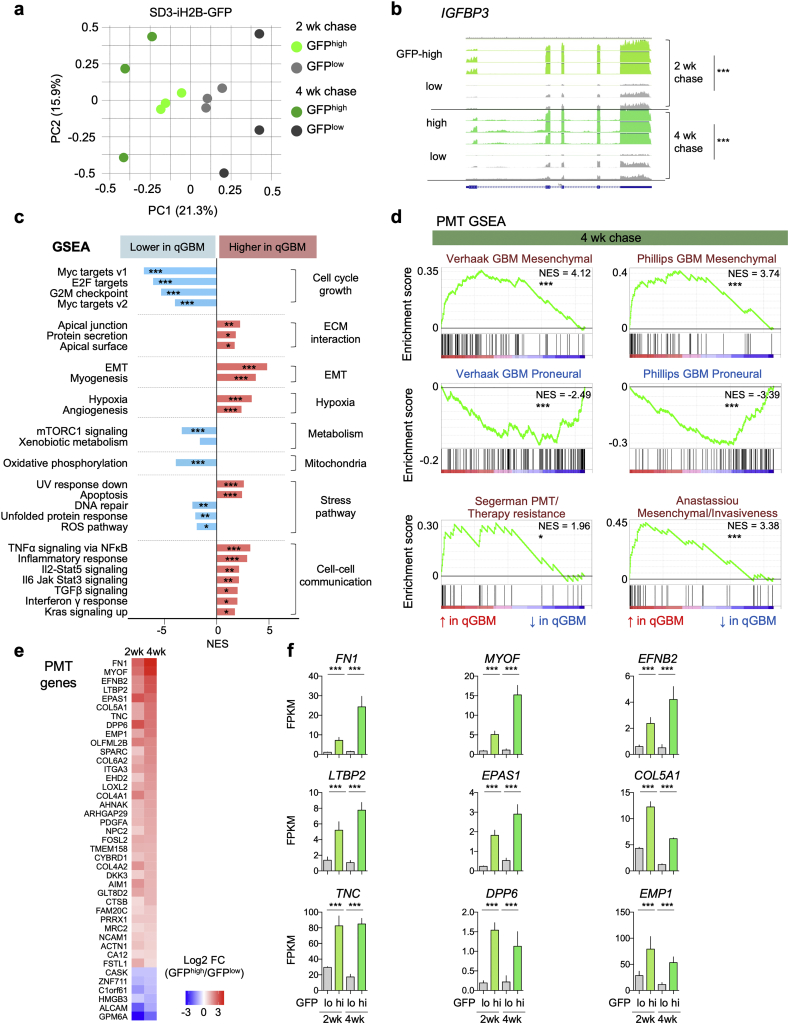
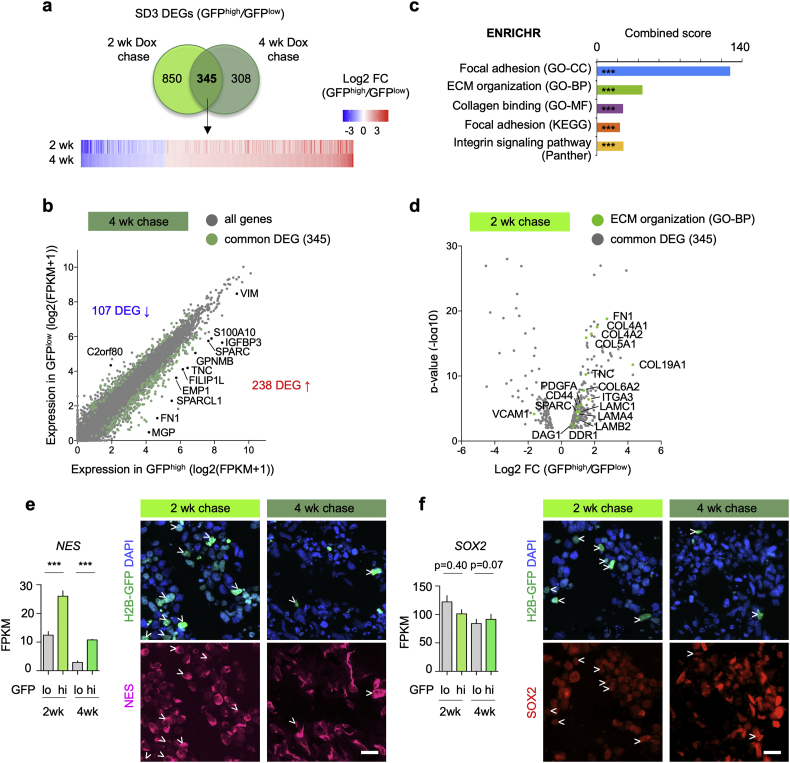
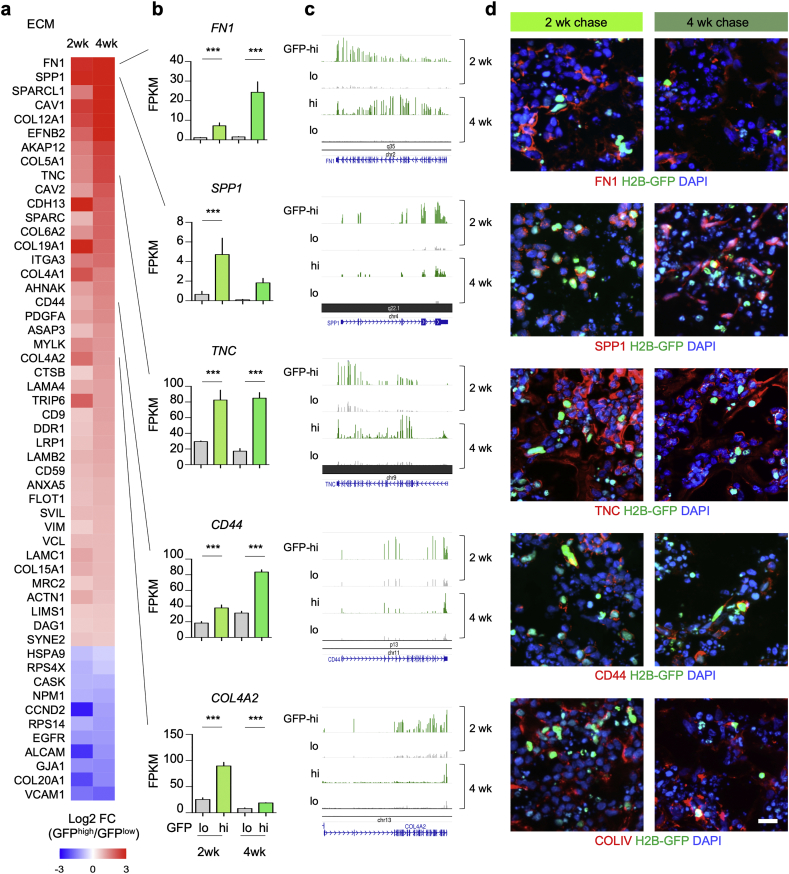
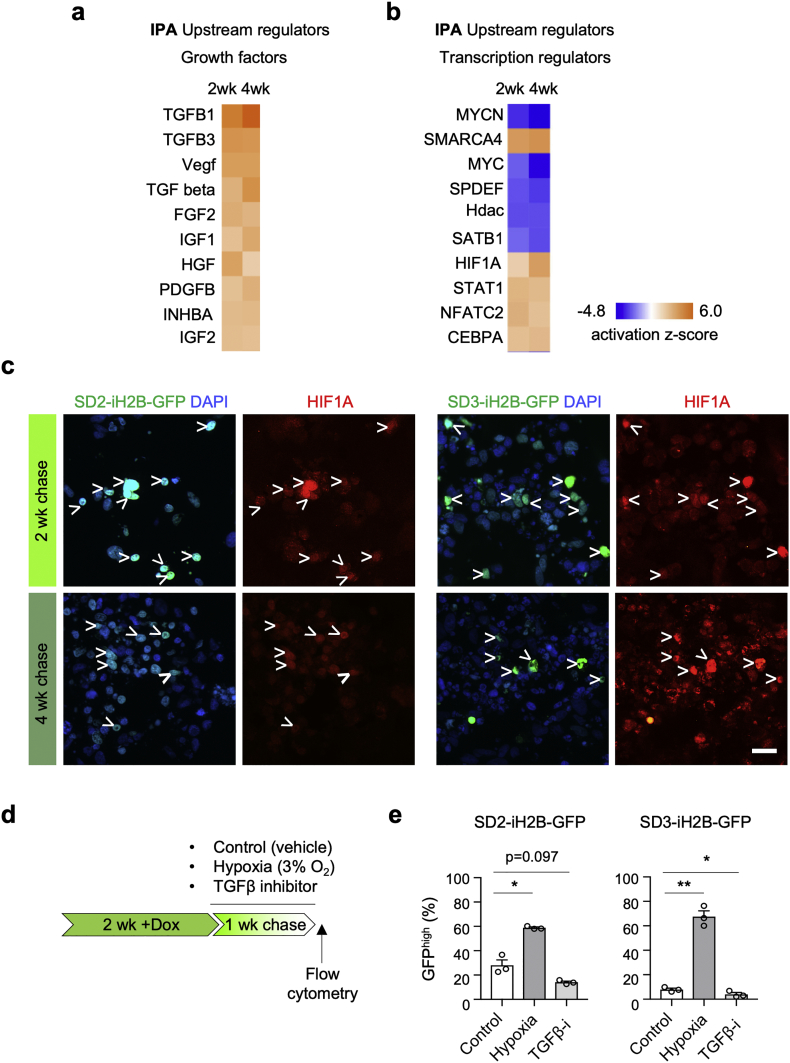
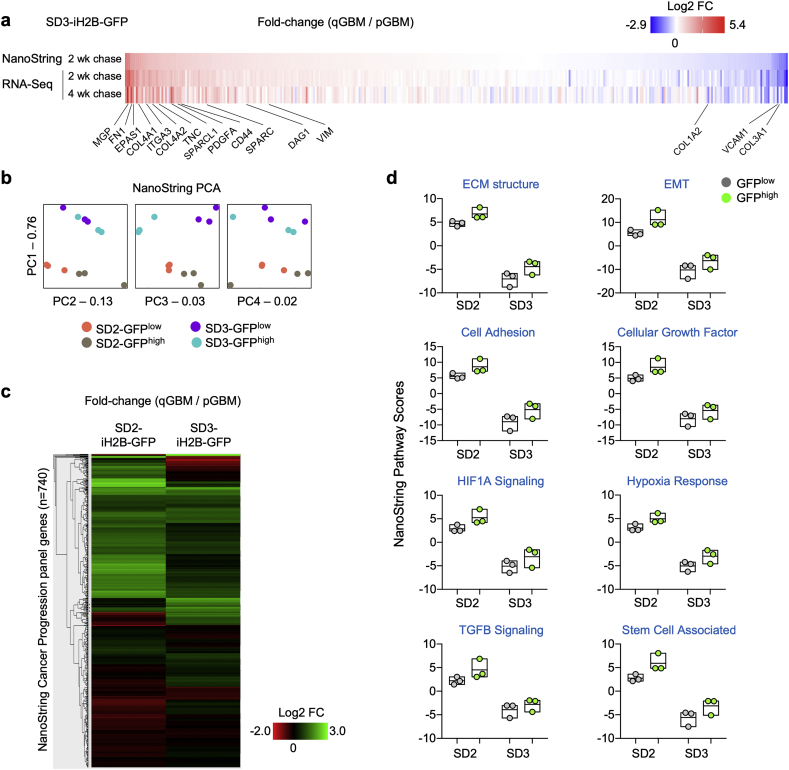
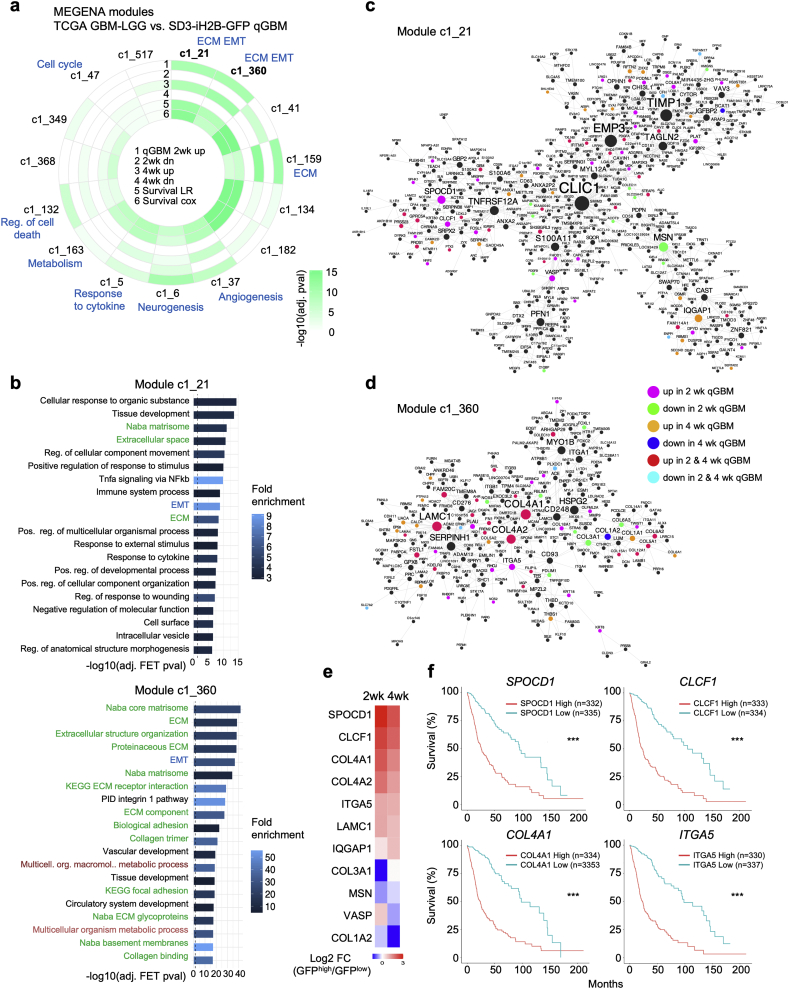
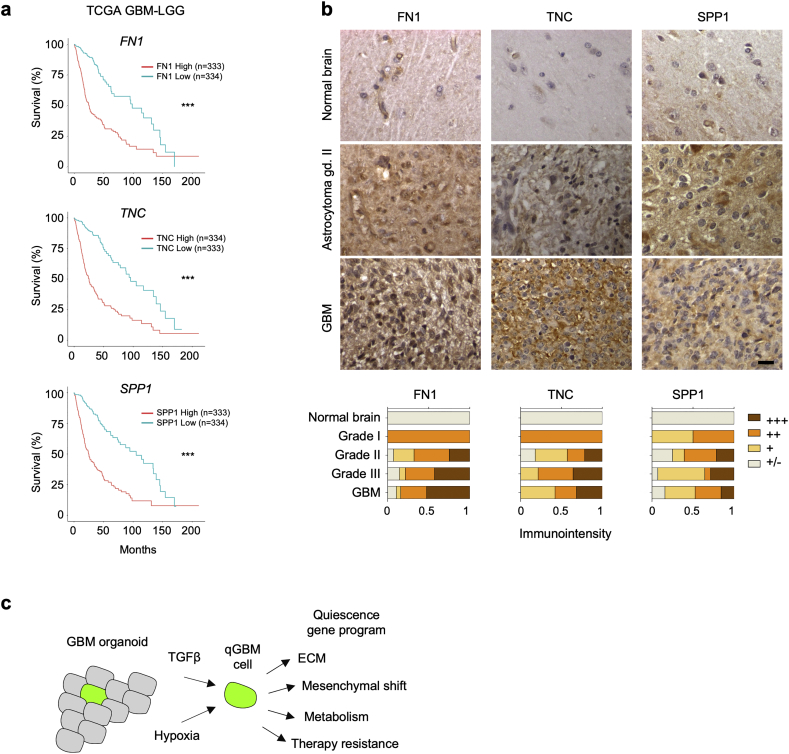
Findings: H2B-GFP-retaining qGBM cells exhibited comparable self-renewal capacity but higher therapy resistance relative to pGBM. Quiescent GBM cells expressed distinct gene programs that affect cell cycle control, metabolic adaptation, and extracellular matrix (ECM) interactions. Transcriptome analysis also revealed a mesenchymal shift in qGBM cells of both proneural and mesenchymal GBM subtypes. Bioinformatic analyses and functional assays in GBM organoids established hypoxia and TGFβ signaling as potential niche factors that promote quiescence in GBM. Finally, network co-expression analysis of TCGA glioma patient data identified gene modules that are enriched for qGBM signatures and also associated with survival rate.
Interpretation: Our in vitro study in 3D GBM organoids supports the presence of a quiescent cell population that displays self-renewal capacity, high therapy resistance, and mesenchymal gene signatures. It also sheds light on how GBM cells may acquire and maintain quiescence through ECM organization and interaction with niche factors such as TGFβ and hypoxia. Our findings provide a starting point for developing strategies to tackle the quiescent population of GBM. FUND: National Institutes of Health (NIH) and Deutsche Forschungsgemeinschaft (DFG).
Keywords: GBM organoid; Glioblastoma; H2B-GFP; Proneural-mesenchymal transition; Stem cell niche; Tumor quiescence.
Copyright © 2019 The Authors. Published by Elsevier B.V. All rights reserved.
Figures
References
-
- Kleinhues P., Barnholtz-Sloan J., Ohgaki H. World Cancer report 2014. International Agency for Research on Cancer; Lyon: 2014. Tumours of the nervous system.
-
- Huse J.T., Holland E.C. Targeting brain cancer: advances in the molecular pathology of malignant glioma and medulloblastoma. Nat Rev Cancer. 2010;10(5):319–331. - PubMed
-
- Visvader J.E. Cells of origin in cancer. Nature. 2011;469:314–322. - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
